Virus Dmitri Ivanovsky tarafından 1892’de keşfedildi (“virus” adı Lat. “virus/zehir” sözcüğünden ödünç alındı; Yunanca köklerden türetilen “bakteriofaj” “bakteri-yiyici” demektir).
Virus DNA ya da RNA olmak üzere genetik gereç taşır, ürer, ve evrimlenir.
Virusların büyük çoğunluğu RNA genomu taşır.
Virus ancak bir örgenliğin (bitki, hayvan, bakteri) dirimli hücresi içinde eşlemlenebilir.
Tüm yaşam biçimlerine bulaşır.
Bulunan milyonlarca virustan yalnızca 6.000 kadarı betimlendi ve 2010 için 74.000’in üzerinde tam virus genom dizisi belirlendi.
Virus genomları dairesel ya da doğrusal olabilir.
En küçük virus genomu (ssDNA circovirus) yalnızca iki protein için kodlayabilir ve büyüklüğü yalnızca iki kilobazdır.
En büyük virus genomu (pandoraviruslarda) yaklaşık iki megabaz büyüklüğündedir 2.500 kadar protein için kodlama yapar.
Konak hücreye bulaşan virus hızla binlerce özdeş eşlemini yapar.
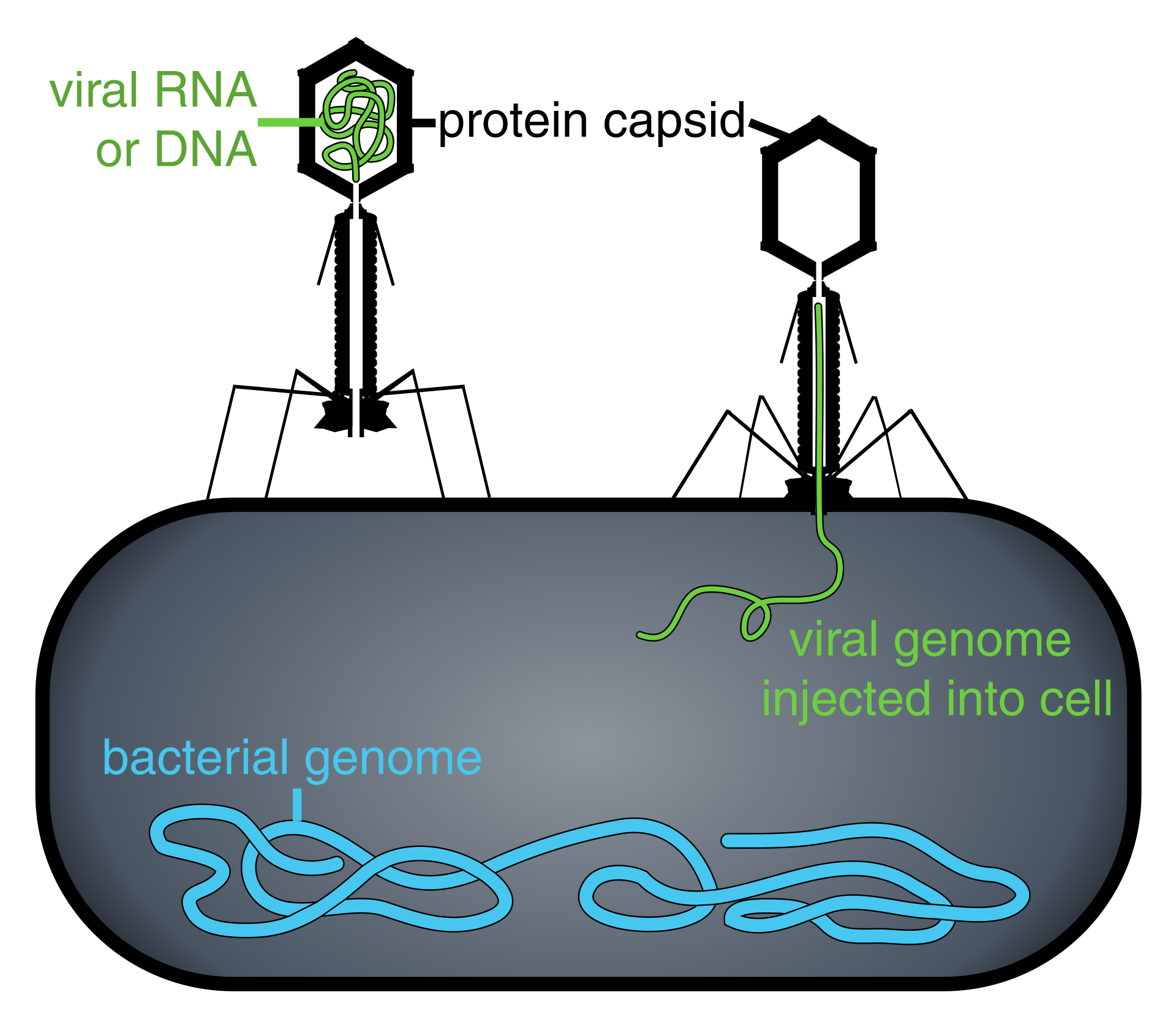
Bulaşma öncesi virus ya da virion (i) uzun DNA ya da RNA molekülleri biçiminde genetik gereçten ve (ii) bir protein kılıf olan kapsidden oluşur; kimi durumlarda (iii) lipidlerden yapılı bir dış zarf bulunur.
Viruslar yalın sarmal ve ikozohedral biçimlerden daha karmaşık yapılara dek çeşitli şekillerde bulunur.
Virionların çoğu ışık mikroskobu ile görülmeyecek denli küçüktür.
Viruslar evrim sürecinde plasmidlerden (hücreler arasında devinen DNA parçaları) ve bakterilerden gelişmiş olabilir.
📹 You Are Made Of Viruses & There’s Nothing You Can Do About It! / Seeker (VİDEO)
📹 You Are Made Of Viruses & There’s Nothing You Can Do About It! / Seeker (LINK)
This CLEAR & SIMPLE tutorial describes the steps of the viral life cycle, including 1) Attachment 2) Entry 3) Replication 4) Biosynthesis 5) Assembly and 6) Release.
📹 Virus DNA in human genome (evolution by infection) / Every Cell A Universe (VİDEO)
📹 Virus DNA in human genome (evolution by infection) / Every Cell A Universe (LINK)
f it wasn't for virus DNA in human genome, humans would NOT exist. The evolution of the mammalian placenta has happened because of endogenous retroviruses. The viruses in human genome have come from infections millions of years ago that have stayed around in our DNA.
The human endogenous retroviruses are on longer functional viruses capable of infecting. Instead, what's left in the human genome are parts of these viruses, specific important genes. These are the critical genes for placental evolution. Placental mammals have adopted these genes in a wonderful way, and turned a tool used by viruses to infect cells into a tool for creating the placenta.
The evolution of the mammalian placenta is not a rare, one time event. Instead, the evolution of the mammalian placenta has occurred over and over. What's even more astounding is that the placenta has evolved in non-mammalian species as well.
We humans need retrovirus genes for our development. We descend from viruses.
📹 What Are Bacteriophages & How Do Phage Viruses Work? / iCONBOY (VİDEO)
📹 What Are Bacteriophages & How Do Phage Viruses Work? / iCONBOY (LINK)
Bacteriophages are viruses that infect bacteria cells, and they are the most abundant biological entities on the planet, estimate at 10^31 in population. I have had the pleasure of working with and researching these phages, and have discovered 3 novel phages on my own. They're great for learning about genomics and bioinformatics because of their small genomes. Either way, I hope you all enjoyed and learned something new!
📹 s The Deadliest Being on Planet Earth — The Bacteriophage / Kurzgesagt (VİDEO)
📹 The Deadliest Being on Planet Earth — The Bacteriophage / Kurzgesagt (LINK)
A war has been raging for billions of years, killing trillions every single day, while we don’t even notice. This war involves the single deadliest being on our planet: The Bacteriophage
There are fossils of viruses, of sorts, preserved in the DNA of the hosts that they’ve infected. Including you. This molecular fossil trail can help us understand where viruses came from, how they evolved and it can even help us tackle the biggest question of all: Are viruses alive?
Explore the lytic and lysogenic viral replication cycles with the Amoeba Sisters! This video also discusses virus structures and why a host is critical for viral reproduction. Expand details for table of contents and further reading suggestions! This updated video replaces our older virus video from 2013.
📹 The Two Viruses That We’ve Had For Millions of Years / PBS Eons (VİDEO)
📹 The Two Viruses That We’ve Had For Millions of Years / PBS Eons (LINK)
There’s one kind of herpesvirus that’s specific to one species of primate, and each virus split off from the herpesvirus family tree when the primate split off from its own tree. But of course, humans are a special kind of primate.
Here are the two studies we discuss in this episode: Wertheim, J. O., Smith, M. D., Smith, D. M., Scheffler, K., & Kosakovsky Pond, S. L. (2014). Evolutionary origins of human herpes simplex viruses 1 and 2. Molecular biology and evolution, 31(9), 2356-2364.
Underdown, S. J., Kumar, K., & Houldcroft, C. (2017). Network analysis of the hominin origin of Herpes Simplex virus 2 from fossil data. Virus evolution, 3(2), vex026.
How the DNA got into the killer machine?
In a bacterium infected by T4, new bacteriophages are assembled in a stepwise process: the shaft builds up, DNA is replicated and the empty pro-head assembles as an empty shell. But how does the DNA get into the prohead?
The initiation of DNA import is not entirely clear, but once DNA, the packaging motor and pro-head interact the DNA is rapidly threaded through a pore in the circular motor at the speed of 2000 basepairs a second.
Once the head is full, the packaging motor cuts the DNA and the complex falls off an expanded head, which has changed in shape. The shaft and long tail fibres are attached to complete the infectious particle. Within 1 hour more than 100 new phages are released from a single infected bacterium which makes the T4 phage one of the most efficient but also fascinating killing machines.
#DNA #bacteriophage #research #medicine #teaching #technology #science
Produced by biolution GmbH 2019
visit please our website: http://visuals.biolution.net/
Model of the T4 Phage
The T4 phage depicted in this video corresponds widely to the current status of research. The model was assembled using structural data available from public databases (see below) and benefitted greatly from the following publications:
Leiman PG, Arisaka F, van Raaij MJ, Kostyuchenko VA, Aksyuk AA, Kanamaru S, Rossmann MG. Morphogenesis of the T4 tail and tail fibers. Virol J. 2010 Dec 3;7:355. doi: 10.1186/1743-422X-7-355.
Eric S. Miller, Elizabeth Kutter, Gisela Mosig, Fumio Arisaka, Takashi Kunisawa, Wolfgang Rüger. Bacteriophage T4 Genome MMBR, Mar. 2003, p. 86–156 Vol. 67, No. 1 doi: 10.1128/MMBR.67.1.86-156.2003
Siying Lin, Tanfis I. Alam, Vishal I. Kottadiel, Carl J. VanGessel, Wei-Chun Tang, Yann R. Chemla and Venigalla B. Rao. Altering the speed of a DNA packaging motor from bacteriophage T4 Nucleic Acids Res. 2017 Nov 2; 45(19): 11437–11448. doi: 10.1093/nar/gkx809
EMDB:EMD-2774, EMD-6079, EMD-6080, EMD-6081, EMD-6082, EMD-6083: Hu B, Margolin W, Molineux IJ, Liu J, Structural remodeling of bacteriophage T4 and host membranes during infection initiation. PNAS 112 e4919-e4928 (2015) doi: 10.1073/pnas.1501064112EMD-8661: Chen Z, Sun L, Zhang Z, Fokine A, Padilla-Sanchez V, Hanein D, Jiang W, Rossmann MG, Rao VB, Cryo-EM structure of the bacteriophage T4 isometric head at 3.3- angstrom resolution and its relevance to the assembly of icosahedral viruses. PNAS 114 E8184-E8193 (2017) doi: 10.1073/pnas.1708483114
These included the following PDB codes:
5VF3 Bacteriophage T4 isometric capsid
3FOH Fitting of gp18M crystal structure into 3D cryo-EM reconstruction of bacteriophage T4 extended tail
5IV5 Cryo-electron microscopy structure of the hexagonal pre-attachment T4 baseplate-tail tube complex
2XGF Structure of the bacteriophage T4 long tail fibre needle-shaped receptor-binding tip
Retrieved from EMDB.org:
PDB consortium, Protein Data Bank: the single global archive for 3D macromolecular structure data (2019). Nucleic acids research Volume 47 p.D520-D528 doi: 10.1093/nar/gky949
Retrieved from rcsb.org
Stephen K Burley, Helen M. Berman, et al. RCSB Protein Data Bank: biological macromolecular structures enabling research and education in fundamental biology, biomedicine, biotechnology and energy (2019) Nucleic Acids Research 47: D464–D474; doi: 10.1093/nar/gky1004.
More than 100 years ago the Franco-Canadian scientist Felix d’Herelle identified bacteriophages! They look like spaceships from another world and are most fearsome killing machines, fortunately they are targeting exclusively bacteria. D’Herelle and his colleague George Eliava immediately realised phages could be weaponised to help patients to fight off bacterial infections. However, in the western hemisphere the control of life-threatening bacterial infections was soon achieved with antibiotics, while phages and viruses became favourite research tools of geneticists. This explains why the bacteriophage T4, which is depicted here, is one of the most intensely studied biological objects. However, phage therapy was pioneered by the Eliava Institute in Georgia, where it has been advanced ever since its discovery.
Phages stick to bacteria with their leg-like fibres, which triggers a change in the protein conformation ramming their shaft into the bacterial hull. The interior of the shaft contains a needle, which punctures the bacterial envelope enabling the phage to inject its genetic material. The phage genome within minutes overcomes the bacterial defences, phage components are assembled within the cell before it ruptures to release a new wave of phages. Several biotech companies are now ready to exploit the deadly potential of phage therapy as a secret weapon to kill superbugs. Phage therapy frequently relies on extensive phage collections.
One of them is maintained with the support of students searching phages, which can be found virtually everywhere. SEA-PHAGES (https://seaphages.org/) is a two-semester, discovery-based undergraduate research course that begins with simple digging in the soil to find new viruses, but progresses through a variety of microbiology techniques and eventually to complex genome annotation and bioinformatic analyses. It has assembled under the supervision of Graham Hatfull's group at the University of Pittsburgh and the Howard Hughes Medical Institute's Science Education division a collection of more than 15.000 phages. In 2019 the Hatfull lab hit the headlines, when they reported in Nature Medicine, that geneticall engineered phages had cured a terminally ill cystic fibrosis patient, who was suffering from multidrug resistant bacterial infections in the lung: https://www.nature.com/articles/s4159...
This is an accurate science animation of the T4 bacteriophage virus that showing how the virus assembles itself after infecting an E. coli cell. These models were generated from data provided by the Michael Rossmann Lab at Purdue University.
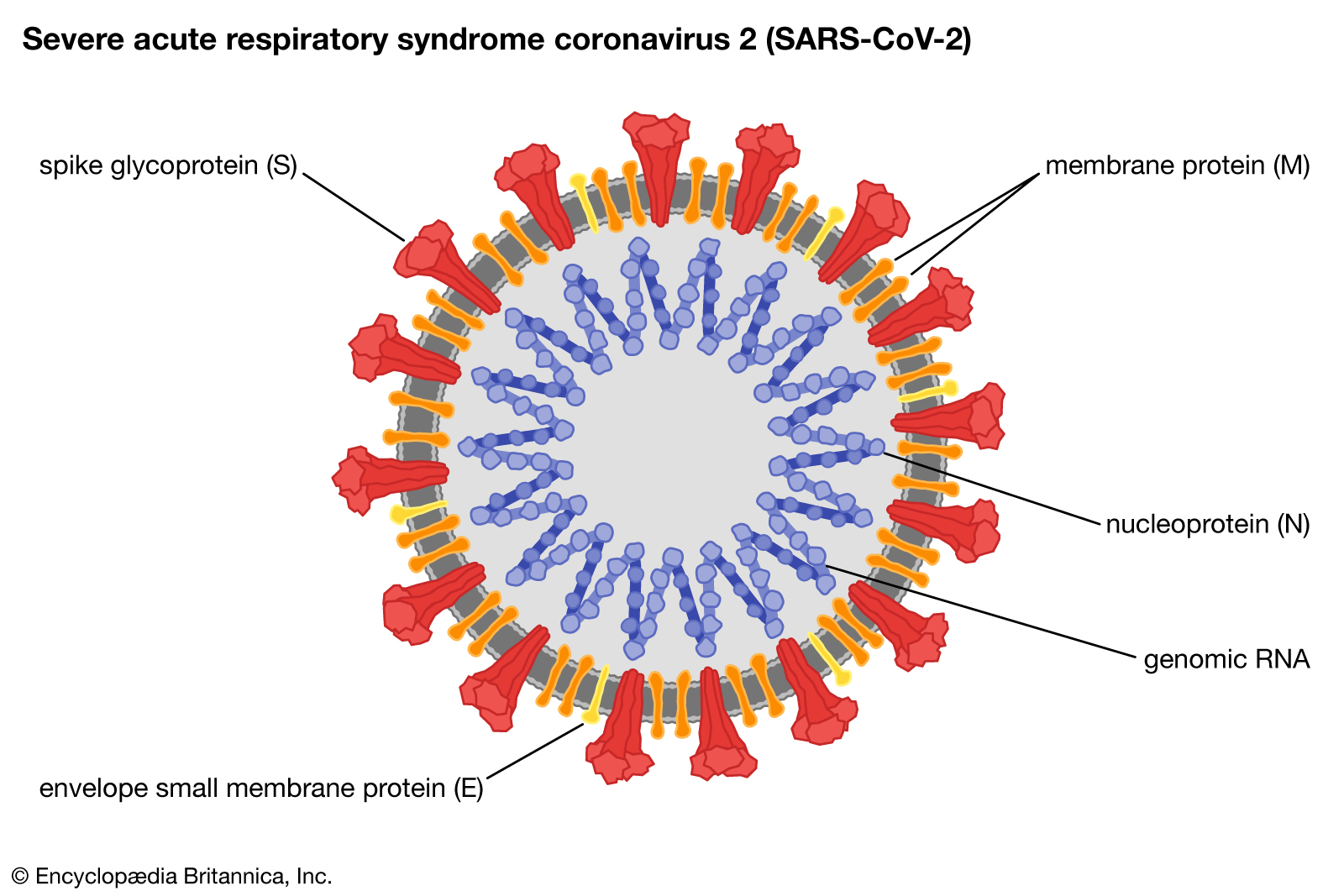
Correction: The video mentions that ACE2 cleaves the spike protein, however a publication published on March 5, 2020 showed that cleavage by the protease TPMSSR2 is required for priming the spike protein for viral entry: https://doi.org/10.1016/j.cell.2020.0...
Thanks to Mikail Dogan, norm1124 and Levent Cavas for pointing this out! We also apologise for referring to the hemagglutinin protein, which is of SARS-CoV. Instead SARS-CoV-2 contains a so called M Protein in the envelope. A correct Figure is here: https://www.ncbi.nlm.nih.gov/pmc/arti...
Thanks to colicinK and Mike Sauder for pointing this out!
📹 What the coronavirus looks like up close? / Vox (VİDEO)
📹 What the coronavirus looks like up close? / Vox (LINK)
When infected, a host cell is forced to rapidly produce thousands of identical copies of the original virus. When not inside an infected cell or in the process of infecting a cell, viruses exist in the form of independent particles, or virions, consisting of: (i) the genetic material, i.e. long molecules of DNA or RNAthat encode the structure of the proteins by which the virus acts; (ii) a protein coat, the capsid, which surrounds and protects the genetic material; and in some cases (iii) an outside envelope of lipids. The shapes of these virus particles range from simple helical and icosahedral forms to more complex structures. Most virus species have virions too small to be seen with an optical microscope as they are one hundredth the size of most bacteria.
The origins of viruses in the evolutionary history of life are unclear: some may have evolved fromplasmids — pieces of DNA that can move between cells — while others may have evolved from bacteria. In evolution, viruses are an important means of horizontal gene transfer, which increases genetic diversity in a way analogous to sexual reproduction. Viruses are considered by some biologiststo be a life form, because they carry genetic material, reproduce, and evolve through natural selection, although they lack the key characteristics such as cell structure that are generally considered necessary criteria for life. Because they possess some but not all such qualities, viruses have been described as "organisms at the edge of life", and as self-replicators.
Viruses spread in many ways. One transmission pathway is through disease-bearing organisms known as vectors: for example, viruses are often transmitted from plant to plant by insects that feed on plant sap, such as aphids; and viruses in animals can be carried by blood-sucking insects. Influenza viruses are spread by coughing and sneezing. Norovirus and rotavirus, common causes of viral gastroenteritis, are transmitted by the faecal–oral route,passed by hand-to-mouth contact or in food or water. The infectious dose of norovirus required to produce infection in humans is less than 100 particles.HIV is one of several viruses transmitted through sexual contact and by exposure to infected blood. The variety of host cells that a virus can infect is called its "host range". This can be narrow, meaning a virus is capable of infecting few species, or broad, meaning it is capable of infecting many.
The word is from the Latin neuter vīrus referring to poison and other noxious liquids, from the same Indo-European base as Sanskritviṣa, Avestanvīša, and ancient Greekἰός (all meaning "poison"), first attested in English in 1398 in John Trevisa's translation of Bartholomeus Anglicus'sDe Proprietatibus Rerum. Virulent, from Latin virulentus (poisonous), dates to c. 1400. A meaning of "agent that causes infectious disease" is first recorded in 1728, long before the discovery of viruses by Dmitri Ivanovskyin 1892. The English plural is viruses (sometimes also vira) whereas the Latin word is a mass noun, which has no classically attested plural (vīra is used in Neo-Latin). The adjective viral dates to 1948. The term virion (plural virions), which dates from 1959, is also used to refer to a single viral particle that is released from the cell and is capable of infecting other cells of the same type.
Louis Pasteur was unable to find a causative agent for rabies and speculated about a pathogen too small to be detected by microscopes. In 1884, the French microbiologistCharles Chamberland invented the Chamberland filter (or Pasteur-Chamberland filter) with pores small enough to remove all bacteria from a solution passed through it. In 1892, the Russian biologist Dmitri Ivanovsky used this filter to study what is now known as the tobacco mosaic virus: crushed leaf extracts from infected tobacco plants remained infectious even after filtration to remove bacteria. Ivanovsky suggested the infection might be caused by a toxin produced by bacteria, but did not pursue the idea. At the time it was thought that all infectious agents could be retained by filters and grown on a nutrient medium — this was part of the germ theory of disease.In 1898, the Dutch microbiologist Martinus Beijerinck repeated the experiments and became convinced that the filtered solution contained a new form of infectious agent. He observed that the agent multiplied only in cells that were dividing, but as his experiments did not show that it was made of particles, he called it a contagium vivum fluidum (soluble living germ) and re-introduced the word virus. Beijerinck maintained that viruses were liquid in nature, a theory later discredited by Wendell Stanley, who proved they were particulate. In the same year Friedrich Loeffler and Paul Frosch passed the first animal virus through a similar filter: aphthovirus, the agent of foot-and-mouth disease.
In the early 20th century, the English bacteriologist Frederick Twort discovered a group of viruses that infect bacteria, now called bacteriophages (or commonly 'phages'), and the French-Canadian microbiologist Félix d'Herelle described viruses that, when added to bacteria on an agar plate, would produce areas of dead bacteria. He accurately diluted a suspension of these viruses and discovered that the highest dilutions (lowest virus concentrations), rather than killing all the bacteria, formed discrete areas of dead organisms. Counting these areas and multiplying by the dilution factor allowed him to calculate the number of viruses in the original suspension. Phages were heralded as a potential treatment for diseases such as typhoid and cholera, but their promise was forgotten with the development of penicillin. The development of bacterial resistance to antibiotics has renewed interest in the therapeutic use of bacteriophages.
By the end of the 19th century, viruses were defined in terms of their infectivity, their ability to pass filters, and their requirement for living hosts. Viruses had been grown only in plants and animals. In 1906, Ross Granville Harrison invented a method for growing tissue in lymph, and, in 1913, E. Steinhardt, C. Israeli, and R. A. Lambert used this method to grow vaccinia virus in fragments of guinea pig corneal tissue. In 1928, H. B. Maitland and M. C. Maitland grew vaccinia virus in suspensions of minced hens' kidneys. Their method was not widely adopted until the 1950s when poliovirus was grown on a large scale for vaccine production.
The first images of viruses were obtained upon the invention of electron microscopy in 1931 by the German engineers Ernst Ruska and Max Knoll. In 1935, American biochemist and virologist Wendell Meredith Stanley examined the tobacco mosaic virus and found it was mostly made of protein. A short time later, this virus was separated into protein and RNA parts. The tobacco mosaic virus was the first to be crystallised and its structure could, therefore, be elucidated in detail. The first X-ray diffraction pictures of the crystallised virus were obtained by Bernal and Fankuchen in 1941. On the basis of her X-ray crystallographic pictures, Rosalind Franklin discovered the full structure of the virus in 1955. In the same year, Heinz Fraenkel-Conrat and Robley Williams showed that purified tobacco mosaic virus RNA and its protein coat can assemble by themselves to form functional viruses, suggesting that this simple mechanism was probably the means through which viruses were created within their host cells.
Viruses are found wherever there is life and have probably existed since living cells first evolved. The origin of viruses is unclear because they do not form fossils, so molecular techniques are used to investigate how they arose. In addition, viral genetic material occasionally integrates into the germline of the host organisms, by which they can be passed on vertically to the offspring of the host for many generations. This provides an invaluable source of information for paleovirologists to trace back ancient viruses that have existed up to millions of years ago. There are three main hypotheses that aim to explain the origins of viruses:
Regressive hypothesis
Viruses may have once been small cells that parasitised larger cells. Over time, genes not required by their parasitism were lost. The bacteria rickettsia and chlamydia are living cells that, like viruses, can reproduce only inside host cells. They lend support to this hypothesis, as their dependence on parasitism is likely to have caused the loss of genes that enabled them to survive outside a cell. This is also called the 'degeneracy hypothesis', or 'reduction hypothesis'.
Cellular origin hypothesis
Some viruses may have evolved from bits of DNA or RNA that "escaped" from the genes of a larger organism. The escaped DNA could have come from plasmids (pieces of naked DNA that can move between cells) or transposons (molecules of DNA that replicate and move around to different positions within the genes of the cell). Once called "jumping genes", transposons are examples of mobile genetic elements and could be the origin of some viruses. They were discovered in maize by Barbara McClintock in 1950. This is sometimes called the 'vagrancy hypothesis', or the 'escape hypothesis'.
Co-evolution hypothesis
This is also called the 'virus-first hypothesis' and proposes that viruses may have evolved from complex molecules of protein and nucleic acid at the same time that cells first appeared on Earth and would have been dependent on cellular life for billions of years. Viroids are molecules of RNA that are not classified as viruses because they lack a protein coat. They have characteristics that are common to several viruses and are often called subviral agents. Viroids are important pathogens of plants. They do not code for proteins but interact with the host cell and use the host machinery for their replication. The hepatitis delta virus of humans has an RNA genome similar to viroids but has a protein coat derived from hepatitis B virus and cannot produce one of its own. It is, therefore, a defective virus. Although hepatitis delta virus genome may replicate independently once inside a host cell, it requires the help of hepatitis B virus to provide a protein coat so that it can be transmitted to new cells. In similar manner, the sputnik virophage is dependent on mimivirus, which infects the protozoan Acanthamoeba castellanii. These viruses, which are dependent on the presence of other virus species in the host cell, are called 'satellites' and may represent evolutionary intermediates of viroids and viruses.
In the past, there were problems with all of these hypotheses: the regressive hypothesis did not explain why even the smallest of cellular parasites do not resemble viruses in any way. The escape hypothesis did not explain the complex capsids and other structures on virus particles. The virus-first hypothesis contravened the definition of viruses in that they require host cells. Viruses are now recognised as ancient and as having origins that pre-date the divergence of life into the three domains. This discovery has led modern virologists to reconsider and re-evaluate these three classical hypotheses.
The evidence for an ancestral world of RNA cells and computer analysis of viral and host DNA sequences are giving a better understanding of the evolutionary relationships between different viruses and may help identify the ancestors of modern viruses. To date, such analyses have not proved which of these hypotheses is correct. It seems unlikely that all currently known viruses have a common ancestor, and viruses have probably arisen numerous times in the past by one or more mechanisms.
Life properties (W)
Scientific opinions differ on whether viruses are a form of life, or organic structures that interact with living organisms. They have been described as "organisms at the edge of life", since they resemble organisms in that they possess genes, evolve by natural selection, and reproduce by creating multiple copies of themselves through self-assembly. Although they have genes, they do not have a cellular structure, which is often seen as the basic unit of life. Viruses do not have their own metabolism, and require a host cell to make new products. They therefore cannot naturally reproduce outside a host cell — although bacterial species such as rickettsia and chlamydia are considered living organisms despite the same limitation. Accepted forms of life use cell division to reproduce, whereas viruses spontaneously assemble within cells. They differ from autonomous growth of crystals as they inherit genetic mutations while being subject to natural selection. Virus self-assembly within host cells has implications for the study of the origin of life, as it lends further credence to the hypothesis that life could have started as self-assembling organic molecules.
Diagram of how a virus capsid can be constructed using multiple copies of just two protein molecules.
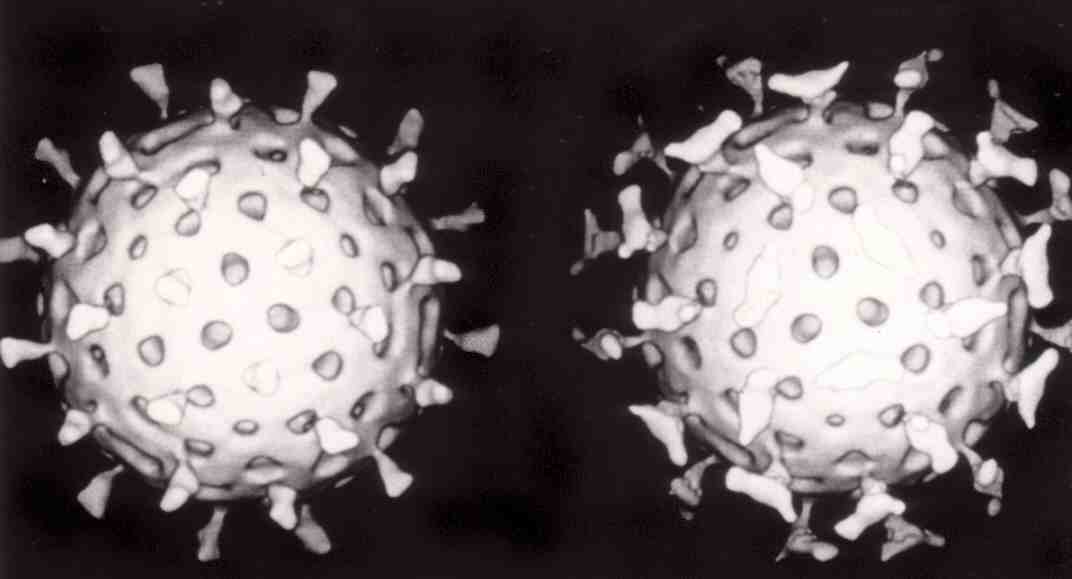
Viruses display a wide diversity of shapes and sizes, called 'morphologies'. In general, viruses are much smaller than bacteria. Most viruses that have been studied have a diameter between 20 and 300 nanometres. Some filoviruses have a total length of up to 1400 nm; their diameters are only about 80 nm. Most viruses cannot be seen with an optical microscope, so scanning and transmission electron microscopes are used to visualise them. To increase the contrast between viruses and the background, electron-dense "stains" are used. These are solutions of salts of heavy metals, such as tungsten, that scatter the electrons from regions covered with the stain. When virions are coated with stain (positive staining), fine detail is obscured. Negative staining overcomes this problem by staining the background only.
A complete virus particle, known as a virion, consists of nucleic acid surrounded by a protective coat of protein called a capsid. These are formed from identical protein subunits called capsomeres. Viruses can have a lipid "envelope" derived from the host cell membrane. The capsid is made from proteins encoded by the viral genome and its shape serves as the basis for morphological distinction. Virally-coded protein subunits will self-assemble to form a capsid, in general requiring the presence of the virus genome. Complex viruses code for proteins that assist in the construction of their capsid. Proteins associated with nucleic acid are known as nucleoproteins, and the association of viral capsid proteins with viral nucleic acid is called a nucleocapsid. The capsid and entire virus structure can be mechanically (physically) probed through atomic force microscopy. In general, there are four main morphological virus types:
These viruses are composed of a single type of capsomere stacked around a central axis to form a helical structure, which may have a central cavity, or tube. This arrangement results in rod-shaped or filamentous virions which can be short and highly rigid, or long and very flexible. The genetic material (typically single-stranded RNA, but ssDNA in some cases) is bound into the protein helix by interactions between the negatively charged nucleic acid and positive charges on the protein. Overall, the length of a helical capsid is related to the length of the nucleic acid contained within it, and the diameter is dependent on the size and arrangement of capsomeres. The well-studied tobacco mosaic virus is an example of a helical virus.
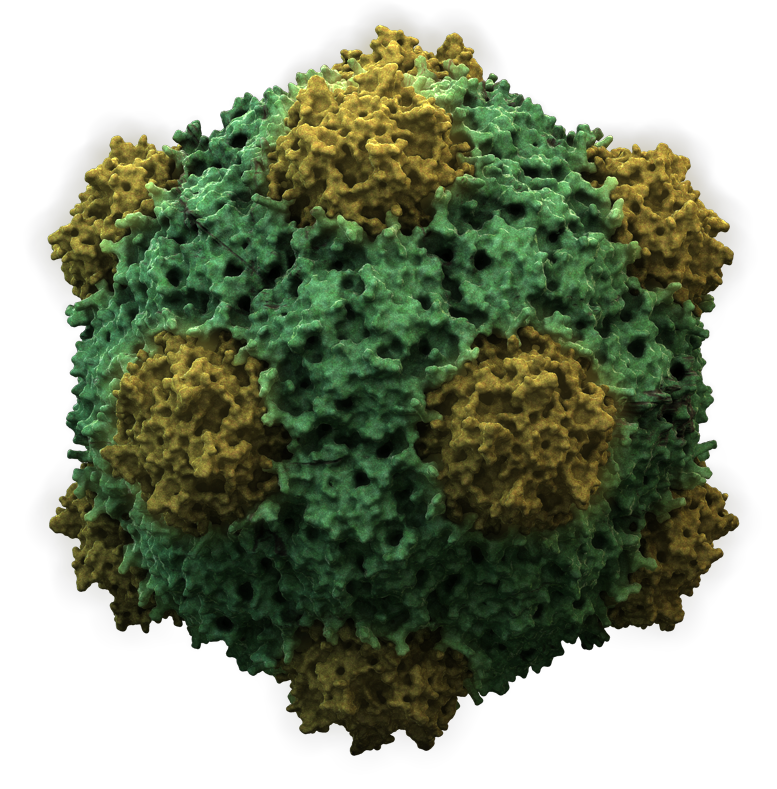
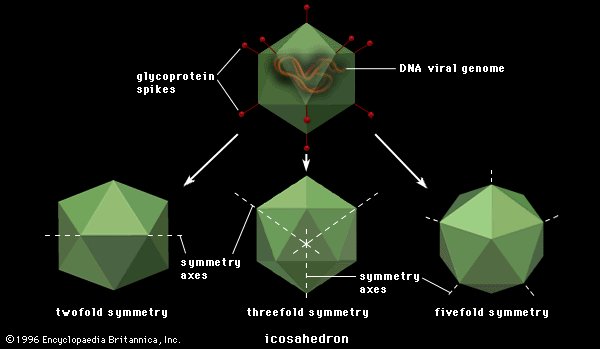
Icosahedral
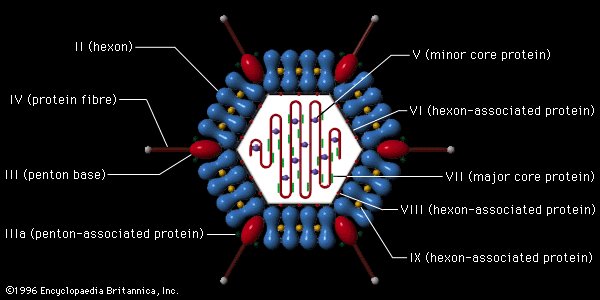
Structure of icosahedral adenovirus. Electron micrograph with an illustration to show shape
Most animal viruses are icosahedral or near-spherical with chiral icosahedral symmetry. A regular icosahedron is the optimum way of forming a closed shell from identical sub-units. The minimum number of identical capsomeres required for each triangular face is 3, which gives 60 for the icosahedron. Many viruses, such as rotavirus, have more than 60 capsomers and appear spherical but they retain this symmetry. To achieve this, the capsomeres at the apices are surrounded by five other capsomeres and are called pentons. Capsomeres on the triangular faces are surrounded by six others and are called hexons. Hexons are in essence flat and pentons, which form the 12 vertices, are curved. The same protein may act as the subunit of both the pentamers and hexamers or they may be composed of different proteins.
Prolate
This is an icosahedron elongated along the fivefold axis and is a common arrangement of the heads of bacteriophages. This structure is composed of a cylinder with a cap at either end.
Envelope
Structure of chickenpox virus. They have a lipid envelope.
Some species of virus envelop themselves in a modified form of one of the cell membranes, either the outer membrane surrounding an infected host cell or internal membranes such as nuclear membrane or endoplasmic reticulum, thus gaining an outer lipid bilayer known as a viral envelope. This membrane is studded with proteins coded for by the viral genome and host genome; the lipid membrane itself and any carbohydrates present originate entirely from the host. The influenza virus and HIV use this strategy. Most enveloped viruses are dependent on the envelope for their infectivity.
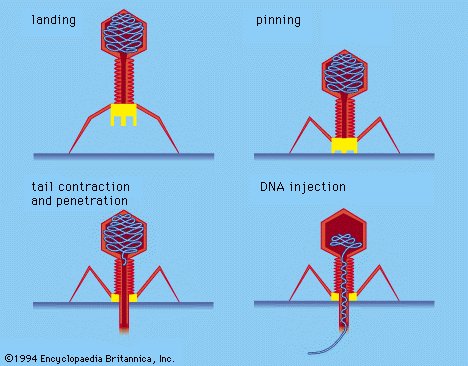
These viruses possess a capsid that is neither purely helical nor purely icosahedral, and that may possess extra structures such as protein tails or a complex outer wall. Some bacteriophages, such as Enterobacteria phage T4, have a complex structure consisting of an icosahedral head bound to a helical tail, which may have a hexagonal base plate with protruding protein tail fibres. This tail structure acts like a molecular syringe, attaching to the bacterial host and then injecting the viral genome into the cell.
The poxviruses are large, complex viruses that have an unusual morphology. The viral genome is associated with proteins within a central disc structure known as a nucleoid. The nucleoid is surrounded by a membrane and two lateral bodies of unknown function. The virus has an outer envelope with a thick layer of protein studded over its surface. The whole virion is slightly pleomorphic, ranging from ovoid to brick-shaped.
Mimivirus is one of the largest characterised viruses, with a capsid diameter of 400 nm. Protein filaments measuring 100 nm project from the surface. The capsid appears hexagonal under an electron microscope, therefore the capsid is probably icosahedral. In 2011, researchers discovered the largest then known virus in samples of water collected from the ocean floor off the coast of Las Cruces, Chile. Provisionally named Megavirus chilensis, it can be seen with a basic optical microscope. In 2013, the Pandoravirus genus was discovered in Chile and Australia, and has genomes about twice as large as Megavirus and Mimivirus. All giant viruses have dsDNA genomes and they are classified into several families: Mimiviridae, Pithoviridae, Pandoraviridae,Phycodnaviridae, and the Mollivirus genus.
Some viruses that infect Archaea have complex structures unrelated to any other form of virus, with a wide variety of unusual shapes, ranging from spindle-shaped structures to viruses that resemble hooked rods, teardrops or even bottles. Other archaeal viruses resemble the tailed bacteriophages, and can have multiple tail structures.
An enormous variety of genomic structures can be seen among viral species; as a group, they contain more structural genomic diversity than plants, animals, archaea, or bacteria. There are millions of different types of viruses, although fewer than 7,000 types have been described in detail. As of September 2015, the NCBI Virus genome database has more than 75,000 complete genome sequences, but there are doubtlessly many more to be discovered.
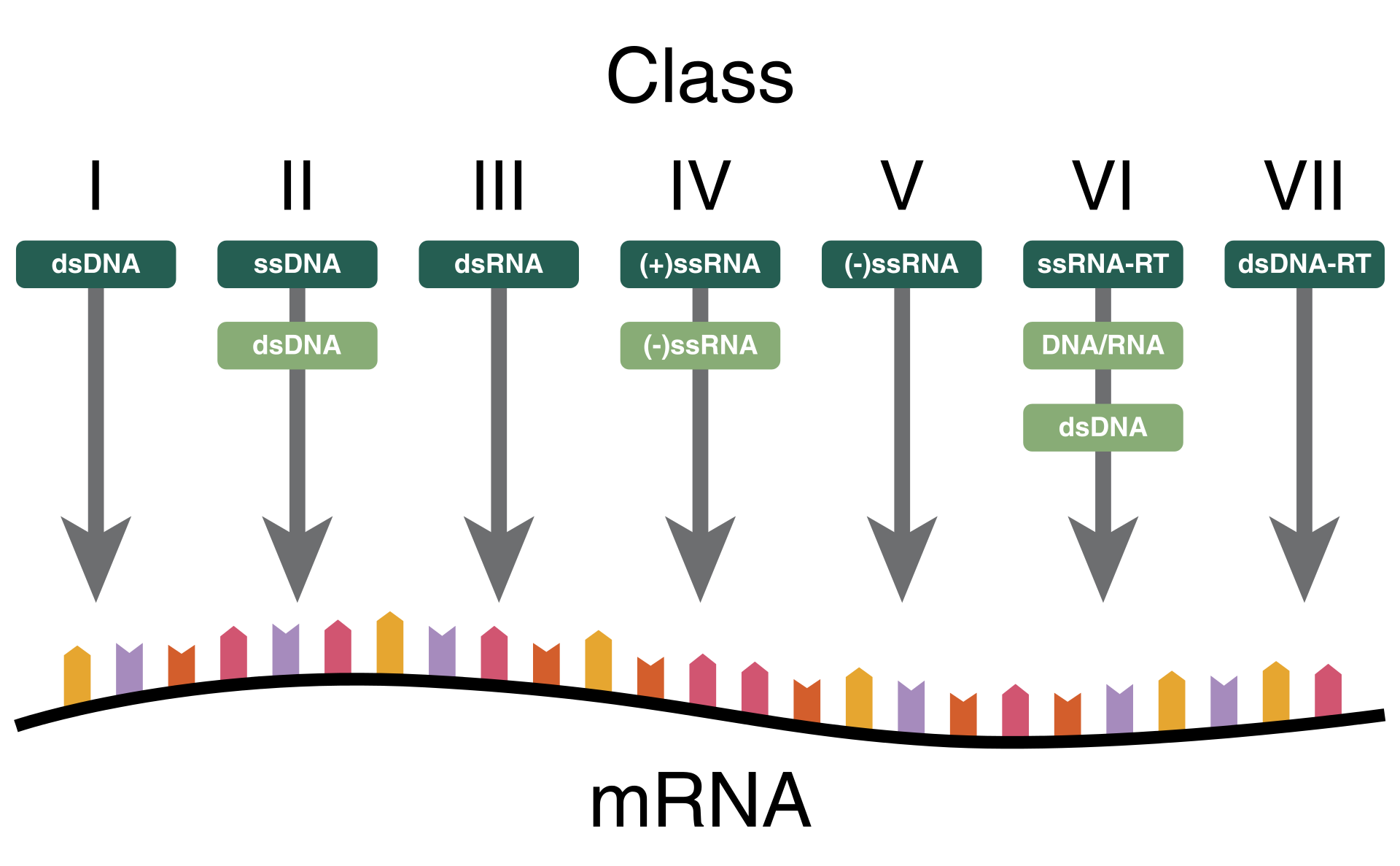
A virus has either a DNA or an RNA genome and is called a DNA virus or an RNA virus, respectively. The vast majority of viruses have RNA genomes. Plant viruses tend to have single-stranded RNA genomes and bacteriophages tend to have double-stranded DNA genomes.
Viral genomes are circular, as in the polyomaviruses, or linear, as in the adenoviruses. The type of nucleic acid is irrelevant to the shape of the genome. Among RNA viruses and certain DNA viruses, the genome is often divided up into separate parts, in which case it is called segmented. For RNA viruses, each segment often codes for only one protein and they are usually found together in one capsid. All segments are not required to be in the same virion for the virus to be infectious, as demonstrated by brome mosaic virus and several other plant viruses.
A viral genome, irrespective of nucleic acid type, is almost always either single-stranded or double-stranded. Single-stranded genomes consist of an unpaired nucleic acid, analogous to one-half of a ladder split down the middle. Double-stranded genomes consist of two complementary paired nucleic acids, analogous to a ladder. The virus particles of some virus families, such as those belonging to the Hepadnaviridae, contain a genome that is partially double-stranded and partially single-stranded.
For most viruses with RNA genomes and some with single-stranded DNA genomes, the single strands are said to be either positive-sense (called the 'plus-strand') or negative-sense (called the 'minus-strand'), depending on if they are complementary to the viral messenger RNA (mRNA). Positive-sense viral RNA is in the same sense as viral mRNA and thus at least a part of it can be immediately translated by the host cell. Negative-sense viral RNA is complementary to mRNA and thus must be converted to positive-sense RNA by an RNA-dependent RNA polymerase before translation. DNA nomenclature for viruses with single-sense genomic ssDNA is similar to RNA nomenclature, in that positive-strand viral ssDNA is identical in sequence to the viral mRNA and is thus a coding strand, while negative-strand viral ssDNA is complementary to the viral mRNA and is thus a template strand. Several types of ssDNA and ssRNA viruses have genomes that are ambisense in that transcription can occur off both strands in a double-stranded replicative intermediate. Examples include geminiviruses, which are ssDNA plant viruses and arenaviruses, which are ssRNA viruses of animals.
Genome size varies greatly between species. The smallest — the ssDNA circoviruses, family Circoviridae — code for only two proteins and have a genome size of only two kilobases; the largest — the pandoraviruses — have genome sizes of around two megabases which code for about 2500 proteins. Virus genes rarely have introns and often are arranged in the genome so that they overlap.
In general, RNA viruses have smaller genome sizes than DNA viruses because of a higher error-rate when replicating, and have a maximum upper size limit. Beyond this, errors when replicating render the virus useless or uncompetitive. To compensate, RNA viruses often have segmented genomes—the genome is split into smaller molecules — thus reducing the chance that an error in a single-component genome will incapacitate the entire genome. In contrast, DNA viruses generally have larger genomes because of the high fidelity of their replication enzymes. Single-strand DNA viruses are an exception to this rule, as mutation rates for these genomes can approach the extreme of the ssRNA virus case.
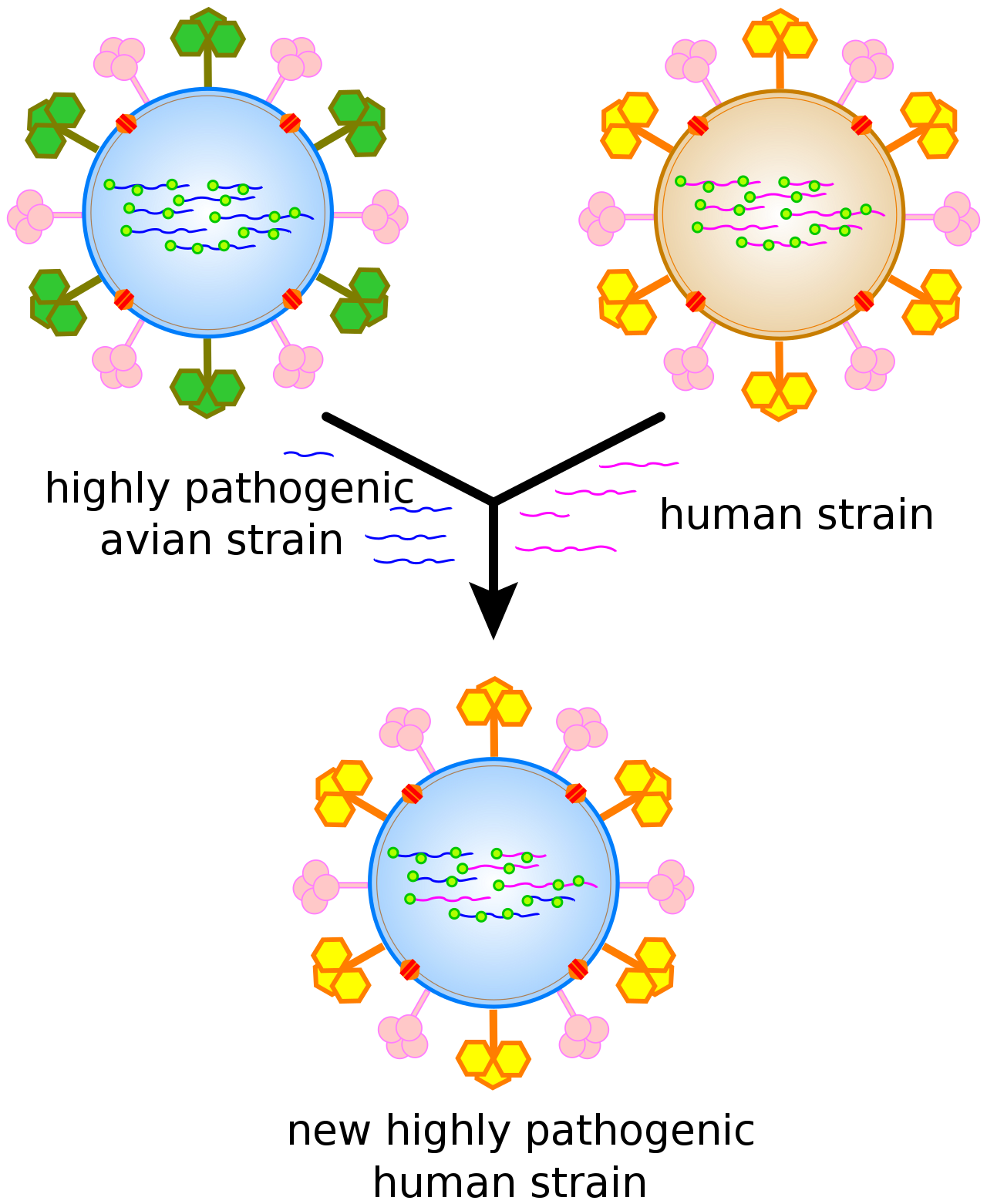
Antigenic shift, or reassortment, can result in novel and highly pathogenic strains of human flu.
Viruses undergo genetic change by several mechanisms. These include a process called antigenic driftwhere individual bases in the DNA or RNA mutate to other bases. Most of these point mutations are "silent" — they do not change the protein that the gene encodes — but others can confer evolutionary advantages such as resistance to antiviral drugs.Antigenic shift occurs when there is a major change in the genome of the virus. This can be a result of recombination or reassortment. When this happens with influenza viruses, pandemics might result. RNA viruses often exist as quasispecies or swarms of viruses of the same species but with slightly different genome nucleoside sequences. Such quasispecies are a prime target for natural selection.
Segmented genomes confer evolutionary advantages; different strains of a virus with a segmented genome can shuffle and combine genes and produce progeny viruses (or offspring) that have unique characteristics. This is called reassortment or 'viral sex'.
Genetic recombination is the process by which a strand of DNA is broken and then joined to the end of a different DNA molecule. This can occur when viruses infect cells simultaneously and studies of viral evolution have shown that recombination has been rampant in the species studied. Recombination is common to both RNA and DNA viruses.
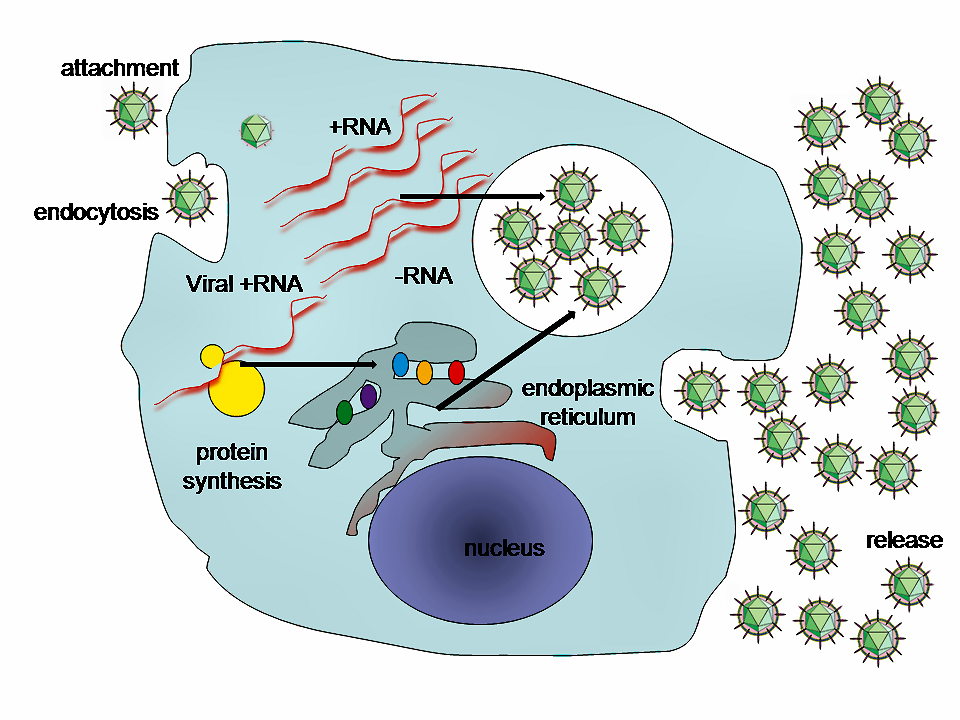
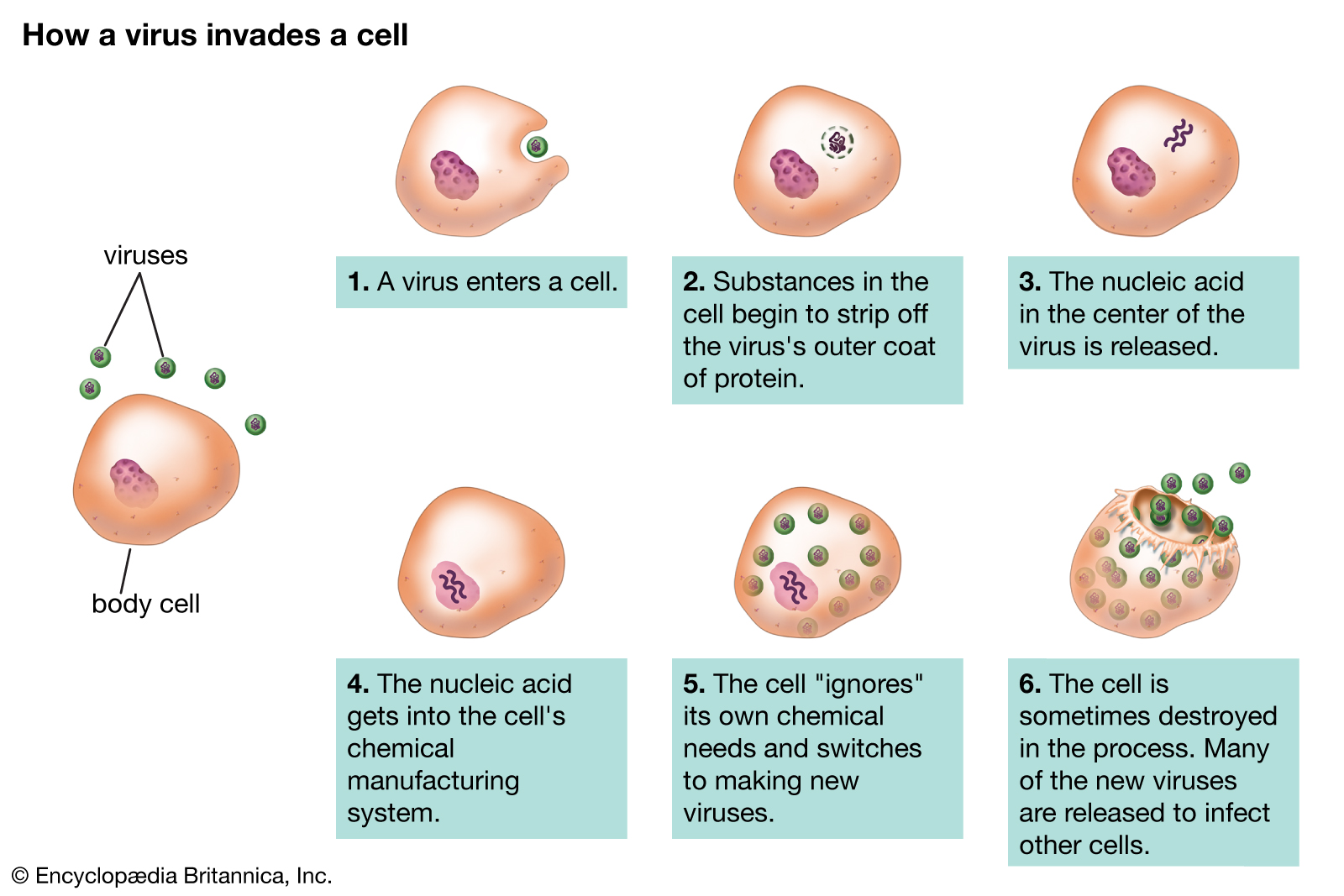
Viral populations do not grow through cell division, because they are acellular. Instead, they use the machinery and metabolism of a host cell to produce multiple copies of themselves, and they assemble in the cell. When infected, the host cell is forced to rapidly produce thousands of identical copies of the original virus.
Their life cycle differs greatly between species, but there are six basic stages in their life cycle:
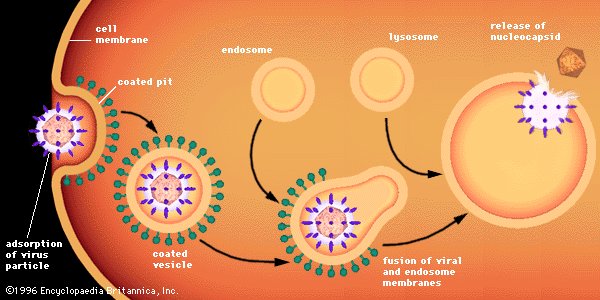
Attachment is a specific binding between viral capsid proteins and specific receptors on the host cellular surface. This specificity determines the host range and type of host cell of a virus. For example, HIV infects a limited range of human leucocytes. This is because its surface protein, gp120, specifically interacts with the CD4 molecule — a chemokine receptor — which is most commonly found on the surface of CD4+T-Cells. This mechanism has evolved to favour those viruses that infect only cells in which they are capable of replication. Attachment to the receptor can induce the viral envelope protein to undergo changes that result in the fusion of viral and cellular membranes, or changes of non-enveloped virus surface proteins that allow the virus to enter.
Penetration follows attachment: Virions enter the host cell through receptor-mediated endocytosis or membrane fusion in a process often known as viral entry. The infection of plant and fungal cells is different from that of animal cells. Plants have a rigid cell wall made of cellulose, and fungi one of chitin, so most viruses can get inside these cells only after trauma to the cell wall. Nearly all plant viruses (such as tobacco mosaic virus) can also move directly from cell to cell, in the form of single-stranded nucleoprotein complexes, through pores called plasmodesmata. Bacteria, like plants, have strong cell walls that a virus must breach to infect the cell. Given that bacterial cell walls are much thinner than plant cell walls due to their much smaller size, some viruses have evolved mechanisms that inject their genome into the bacterial cell across the cell wall, while the viral capsid remains outside.
Uncoating is a process in which the viral capsid is removed: This may be by degradation by viral enzymes or host enzymes or by simple dissociation; the end-result is the releasing of the viral genomic nucleic acid.
A typical virus replication cycle
Replication of viruses involves primarily multiplication of the genome. Replication involves synthesis of viral messenger RNA (mRNA) from "early" genes (with exceptions for positive sense RNA viruses), viral protein synthesis, possible assembly of viral proteins, then viral genome replication mediated by early or regulatory protein expression. This may be followed, for complex viruses with larger genomes, by one or more further rounds of mRNA synthesis: "late" gene expression is, in general, of structural or virion proteins.
Assembly – Following the structure-mediated self-assembly of the virus particles, some modification of the proteins often occurs. In viruses such as HIV, this modification (sometimes called maturation) occurs after the virus has been released from the host cell.
Release – Viruses can be released from the host cell by lysis, a process that kills the cell by bursting its membrane and cell wall if present: this is a feature of many bacterial and some animal viruses. Some viruses undergo a lysogenic cycle where the viral genome is incorporated by genetic recombination into a specific place in the host's chromosome. The viral genome is then known as a “provirus” or, in the case of bacteriophages a “prophage”. Whenever the host divides, the viral genome is also replicated. The viral genome is mostly silent within the host. At some point, the provirus or prophage may give rise to active virus, which may lyse the host cells. Enveloped viruses (e.g., HIV) typically are released from the host cell by budding. During this process the virus acquires its envelope, which is a modified piece of the host's plasma or other, internal membrane.
Some bacteriophages inject their genomes into bacterial cells (not to scale).
The genetic material within virus particles, and the method by which the material is replicated, varies considerably between different types of viruses.
DNA viruses
The genome replication of most DNA viruses takes place in the cell's nucleus. If the cell has the appropriate receptor on its surface, these viruses enter the cell either by direct fusion with the cell membrane (e.g., herpesviruses) or—more usually—by receptor-mediated endocytosis. Most DNA viruses are entirely dependent on the host cell's DNA and RNA synthesising machinery, and RNA processing machinery. Viruses with larger genomes may encode much of this machinery themselves. In eukaryotes the viral genome must cross the cell's nuclear membrane to access this machinery, while in bacteria it need only enter the cell.
RNA viruses
Replication of RNA viruses usually takes place in the cytoplasm. RNA viruses can be placed into four different groups depending on their modes of replication. The polarity (whether or not it can be used directly by ribosomes to make proteins) of single-stranded RNA viruses largely determines the replicative mechanism; the other major criterion is whether the genetic material is single-stranded or double-stranded. All RNA viruses use their own RNA replicase enzymes to create copies of their genomes.
Reverse transcribing viruses
Reverse transcribing viruses have ssRNA (Retroviridae, Metaviridae, Pseudoviridae) or dsDNA (Caulimoviridae, and Hepadnaviridae) in their particles. Reverse transcribing viruses with RNA genomes (retroviruses) use a DNA intermediate to replicate, whereas those with DNA genomes (pararetroviruses) use an RNA intermediate during genome replication. Both types use a reverse transcriptase, or RNA-dependent DNA polymerase enzyme, to carry out the nucleic acid conversion. Retroviruses integrate the DNA produced by reverse transcription into the host genome as a provirus as a part of the replication process; pararetroviruses do not, although integrated genome copies of especially plant pararetroviruses can give rise to infectious virus. They are susceptible to antiviral drugs that inhibit the reverse transcriptase enzyme, e.g. zidovudine and lamivudine. An example of the first type is HIV, which is a retrovirus. Examples of the second type are the Hepadnaviridae, which includes Hepatitis B virus.
Cytopathic effects on the host cell
Cytopathic effects on the host cell (W)
The range of structural and biochemical effects that viruses have on the host cell is extensive. These are called 'cytopathic effects'. Most virus infections eventually result in the death of the host cell. The causes of death include cell lysis, alterations to the cell's surface membrane and apoptosis. Often cell death is caused by cessation of its normal activities because of suppression by virus-specific proteins, not all of which are components of the virus particle. The distinction between cytopathic and harmless is gradual. Some viruses, such as Epstein–Barr virus, can cause cells to proliferate without causing malignancy, while others, such as papillomaviruses, are established causes of cancer.
Some viruses cause no apparent changes to the infected cell. Cells in which the virus is latent and inactive show few signs of infection and often function normally. This causes persistent infections and the virus is often dormant for many months or years. This is often the case with herpes viruses.
Viruses are by far the most abundant biological entities on Earth and they outnumber all the others put together. They infect all types of cellular life including animals, plants, bacteria and fungi. Different types of viruses can infect only a limited range of hosts and many are species-specific. Some, such as smallpox virus for example, can infect only one species—in this case humans, and are said to have a narrow host range. Other viruses, such as rabies virus, can infect different species of mammals and are said to have a broad range. The viruses that infect plants are harmless to animals, and most viruses that infect other animals are harmless to humans. The host range of some bacteriophages is limited to a single strain of bacteria and they can be used to trace the source of outbreaks of infections by a method called phage typing. The complete set of viruses in an organism or habitat is called the virome; for example, all human viruses constitute the human virome.
Classification
Classification
Classification (W)
Classification seeks to describe the diversity of viruses by naming and grouping them on the basis of similarities. In 1962, André Lwoff, Robert Horne, and Paul Tournier were the first to develop a means of virus classification, based on the Linnaean hierarchical system. This system based classification on phylum,class,order,family,genus, and species. Viruses were grouped according to their shared properties (not those of their hosts) and the type of nucleic acid forming their genomes. In 1966, the International Committee on Taxonomy of Viruses (ICTV) was formed. The system proposed by Lwoff, Horne and Tournier was initially not accepted by the ICTV because the small genome size of viruses and their high rate of mutation made it difficult to determine their ancestry beyond order. As such, the Baltimore classification system has come to be used to supplement the more traditional hierarchy. Starting in 2018, the ICTV began to acknowledge deeper evolutionary relationships between viruses that have been discovered over time and adopted a 15-rank classification system ranging from realm to species.
The ICTV developed the current classification system and wrote guidelines that put a greater weight on certain virus properties to maintain family uniformity. A unified taxonomy (a universal system for classifying viruses) has been established. Only a small part of the total diversity of viruses has been studied. As of 2019, 4 realms, 9 kingdoms, 16 phyla, 2 subphyla, 36 classes, 55 orders, 8 suborders, 168 families, 103 subfamilies,1,421 genera, 68 subgenera, and 6,589 species of viruses have been defined by the ICTV.
The general taxonomic structure of taxon ranges and the suffixes used in taxonomic names are shown hereafter. As of 2019, the ranks of subrealm, subkingdom, and subclass are unused, whereas all other ranks are in use.
The Nobel Prize-winning biologist David Baltimore devised the Baltimore classification system. The ICTV classification system is used in conjunction with the Baltimore classification system in modern virus classification.
The Baltimore classification of viruses is based on the mechanism of mRNA production. Viruses must generate mRNAs from their genomes to produce proteins and replicate themselves, but different mechanisms are used to achieve this in each virus family. Viral genomes may be single-stranded (ss) or double-stranded (ds), RNA or DNA, and may or may not use reverse transcriptase (RT). In addition, ssRNA viruses may be either sense (+) or antisense (−). This classification places viruses into seven groups:
Viruses have different mechanisms by which they produce disease in an organism, which depends largely on the viral species. Mechanisms at the cellular level primarily include cell lysis, the breaking open and subsequent death of the cell. In multicellular organisms, if enough cells die, the whole organism will start to suffer the effects. Although viruses cause disruption of healthy homeostasis, resulting in disease, they may exist relatively harmlessly within an organism. An example would include the ability of the herpes simplex virus, which causes cold sores, to remain in a dormant state within the human body. This is called latency and is a characteristic of the herpes viruses, including Epstein–Barr virus, which causes glandular fever, and varicella zoster virus, which causes chickenpox and shingles. Most people have been infected with at least one of these types of herpes virus. These latent viruses might sometimes be beneficial, as the presence of the virus can increase immunity against bacterial pathogens, such as Yersinia pestis.
Some viruses can cause lifelong or chronic infections, where the viruses continue to replicate in the body despite the host's defence mechanisms. This is common in hepatitis B virus and hepatitis C virus infections. People chronically infected are known as carriers, as they serve as reservoirs of infectious virus. In populations with a high proportion of carriers, the disease is said to be endemic.
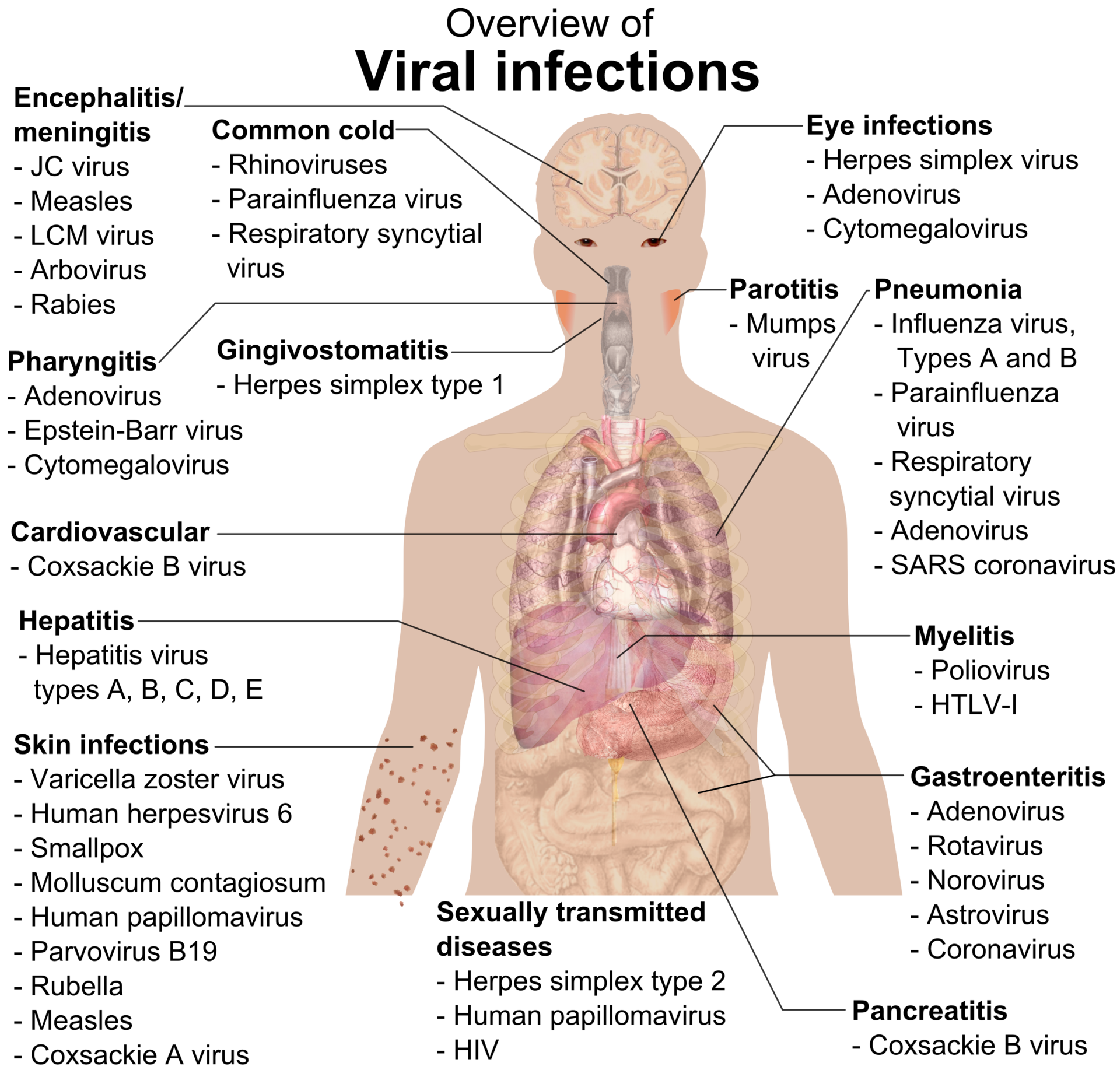
Overview of the main types of viral infection and the most notable species involved.
Viral epidemiology is the branch of medical science that deals with the transmission and control of virus infections in humans. Transmission of viruses can be vertical, which means from mother to child, or horizontal, which means from person to person. Examples of vertical transmission include hepatitis B virus and HIV, where the baby is born already infected with the virus. Another, more rare, example is the varicella zoster virus, which, although causing relatively mild infections in children and adults, can be fatal to the foetus and newborn baby.
Horizontal transmission is the most common mechanism of spread of viruses in populations. Horizontal transmission can occur when body fluids are exchanged during sexual activity, by exchange of saliva or when contaminated food or water is ingested. It can also occur when aerosols containing viruses are inhaled or by insect vectors such as when infected mosquitoes penetrate the skin of a host. Most types of viruses are restricted to just one or two of these mechanisms and they are referred to as "respiratory viruses" or "enteric viruses" and so forth. The rate or speed of transmission of viral infections depends on factors that include population density, the number of susceptible individuals, (i.e., those not immune), the quality of healthcare and the weather.
Epidemiology is used to break the chain of infection in populations during outbreaks of viral diseases. Control measures are used that are based on knowledge of how the virus is transmitted. It is important to find the source, or sources, of the outbreak and to identify the virus. Once the virus has been identified, the chain of transmission can sometimes be broken by vaccines. When vaccines are not available, sanitation and disinfection can be effective. Often, infected people are isolated from the rest of the community, and those that have been exposed to the virus are placed in quarantine. To control the outbreak of foot-and-mouth disease in cattle in Britain in 2001, thousands of cattle were slaughtered. Most viral infections of humans and other animals have incubation periods during which the infection causes no signs or symptoms. Incubation periods for viral diseases range from a few days to weeks, but are known for most infections. Somewhat overlapping, but mainly following the incubation period, there is a period of communicability—a time when an infected individual or animal is contagious and can infect another person or animal. This, too, is known for many viral infections, and knowledge of the length of both periods is important in the control of outbreaks. When outbreaks cause an unusually high proportion of cases in a population, community, or region, they are called epidemics. If outbreaks spread worldwide, they are called pandemics
A pandemic is a worldwide epidemic. The 1918 flu pandemic, which lasted until 1919, was a category 5 influenza pandemic caused by an unusually severe and deadly influenza A virus. The victims were often healthy young adults, in contrast to most influenza outbreaks, which predominantly affect juvenile, elderly, or otherwise-weakened patients. Older estimates say it killed 40–50 million people, while more recent research suggests that it may have killed as many as 100 million people, or 5% of the world's population in 1918.
Although viral pandemics are rare events, HIV—which evolved from viruses found in monkeys and chimpanzees—has been pandemic since at least the 1980s. During the 20th century there were four pandemics caused by influenza virus and those that occurred in 1918, 1957 and 1968 were severe. Most researchers believe that HIV originated in sub-Saharan Africa during the 20th century; it is now a pandemic, with an estimated 37.9 million people now living with the disease worldwide. There were about 770,000 deaths from AIDS in 2018. The Joint United Nations Programme on HIV/AIDS (UNAIDS) and the World Health Organization (WHO) estimate that AIDS has killed more than 25 million people since it was first recognised on 5 June 1981, making it one of the most destructive epidemics in recorded history. In 2007 there were 2.7 million new HIV infections and 2 million HIV-related deaths.
With the exception of smallpox, most pandemics are caused by newly evolved viruses. These "emergent" viruses are usually mutants of less harmful viruses that have circulated previously either in humans or other animals.
Severe acute respiratory syndrome (SARS) and Middle East respiratory syndrome (MERS) are caused by new types of coronaviruses. Other coronaviruses are known to cause mild infections in humans, so the virulence and rapid spread of SARS infections—that by July 2003 had caused around 8,000 cases and 800 deaths—was unexpected and most countries were not prepared.
A related coronavirus emerged in Wuhan, China in November 2019 and spread rapidly around the world. Thought to have originated in bats and subsequently named severe acute respiratory syndrome coronavirus 2, infections with the virus caused a pandemic in 2020. Unprecedented restrictions in peacetime have been placed on international travel, and curfews imposed in several major cities worldwide.
The body's first line of defence against viruses is the innate immune system. This comprises cells and other mechanisms that defend the host from infection in a non-specific manner. This means that the cells of the innate system recognise, and respond to, pathogens in a generic way, but, unlike the adaptive immune system, it does not confer long-lasting or protective immunity to the host.
Two rotaviruses: the one on the right is coated with antibodies that prevent its attachment to cells and infecting them.
RNA interference is an important innate defence against viruses. Many viruses have a replication strategy that involves double-stranded RNA (dsRNA). When such a virus infects a cell, it releases its RNA molecule or molecules, which immediately bind to a protein complex called a dicer that cuts the RNA into smaller pieces. A biochemical pathway—the RISC complex—is activated, which ensures cell survival by degrading the viral mRNA. Rotaviruses have evolved to avoid this defence mechanism by not uncoating fully inside the cell, and releasing newly produced mRNA through pores in the particle's inner capsid. Their genomic dsRNA remains protected inside the core of the virion.
When the adaptive immune system of a vertebrate encounters a virus, it produces specific antibodies that bind to the virus and often render it non-infectious. This is called humoral immunity. Two types of antibodies are important. The first, called IgM, is highly effective at neutralising viruses but is produced by the cells of the immune system only for a few weeks. The second, called IgG, is produced indefinitely. The presence of IgM in the blood of the host is used to test for acute infection, whereas IgG indicates an infection sometime in the past. IgG antibody is measured when tests for immunity are carried out.
Antibodies can continue to be an effective defence mechanism even after viruses have managed to gain entry to the host cell. A protein that is in cells, called TRIM21, can attach to the antibodies on the surface of the virus particle. This primes the subsequent destruction of the virus by the enzymes of the cell's proteosome system.
A second defence of vertebrates against viruses is called cell-mediated immunity and involves immune cells known as T cells. The body's cells constantly display short fragments of their proteins on the cell's surface, and, if a T cell recognises a suspicious viral fragment there, the host cell is destroyed by 'killer T' cells and the virus-specific T-cells proliferate. Cells such as the macrophage are specialists at this antigen presentation. The production of interferon is an important host defence mechanism. This is a hormone produced by the body when viruses are present. Its role in immunity is complex; it eventually stops the viruses from reproducing by killing the infected cell and its close neighbours.
Not all virus infections produce a protective immune response in this way. HIV evades the immune system by constantly changing the amino acid sequence of the proteins on the surface of the virion. This is known as "escape mutation" as the viral epitopes escape recognition by the host immune response. These persistent viruses evade immune control by sequestration, blockade of antigen presentation,cytokine resistance, evasion of natural killer cell activities, escape from apoptosis, and antigenic shift. Other viruses, called 'neurotropic viruses', are disseminated by neural spread where the immune system may be unable to reach them.
Because viruses use vital metabolic pathways within host cells to replicate, they are difficult to eliminate without using drugs that cause toxic effects to host cells in general. The most effective medical approaches to viral diseases are vaccinations to provide immunity to infection, and antiviral drugs that selectively interfere with viral replication.
Vaccination is a cheap and effective way of preventing infections by viruses. Vaccines were used to prevent viral infections long before the discovery of the actual viruses. Their use has resulted in a dramatic decline in morbidity (illness) and mortality (death) associated with viral infections such as polio,measles,mumps and rubella.Smallpox infections have been eradicated. Vaccines are available to prevent over thirteen viral infections of humans, and more are used to prevent viral infections of animals. Vaccines can consist of live-attenuated or killed viruses, or viral proteins (antigens). Live vaccines contain weakened forms of the virus, which do not cause the disease but, nonetheless, confer immunity. Such viruses are called attenuated. Live vaccines can be dangerous when given to people with a weak immunity (who are described as immunocompromised), because in these people, the weakened virus can cause the original disease. Biotechnology and genetic engineering techniques are used to produce subunit vaccines. These vaccines use only the capsid proteins of the virus. Hepatitis B vaccine is an example of this type of vaccine. Subunit vaccines are safe for immunocompromised patients because they cannot cause the disease. The yellow fever virus vaccine, a live-attenuated strain called 17D, is probably the safest and most effective vaccine ever generated
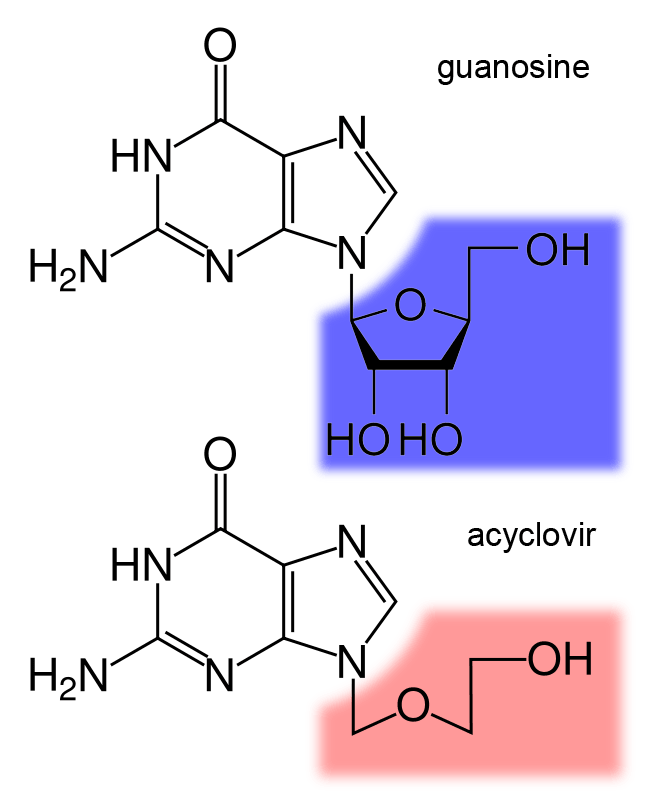
The structure of the DNA base guanosine and the antiviral drug acyclovir
Antiviral drugs are often nucleoside analogues (fake DNA building-blocks), which viruses mistakenly incorporate into their genomes during replication. The life-cycle of the virus is then halted because the newly synthesised DNA is inactive. This is because these analogues lack the hydroxyl groups, which, along with phosphorus atoms, link together to form the strong "backbone" of the DNA molecule. This is called DNA chain termination. Examples of nucleoside analogues are aciclovir for Herpes simplex virus infections and lamivudine for HIV and hepatitis B virus infections. Aciclovir is one of the oldest and most frequently prescribed antiviral drugs. Other antiviral drugs in use target different stages of the viral life cycle. HIV is dependent on a proteolytic enzyme called the HIV-1 protease for it to become fully infectious. There is a large class of drugs called protease inhibitors that inactivate this enzyme.
Hepatitis C is caused by an RNA virus. In 80% of people infected, the disease is chronic, and without treatment, they are infected for the remainder of their lives. There is now an effective treatment that uses the nucleoside analogue drug ribavirin combined with interferon. The treatment of chronic carriers of the hepatitis B virus by using a similar strategy using lamivudine has been developed.
Infection in other species
Infection in other species
Infection in other species (W)
Viruses infect all cellular life and, although viruses occur universally, each cellular species has its own specific range that often infect only that species. Some viruses, called satellites, can replicate only within cells that have already been infected by another virus.
Viruses are important pathogens of livestock. Diseases such as foot-and-mouth disease and bluetongue are caused by viruses. Companion animals such as cats, dogs, and horses, if not vaccinated, are susceptible to serious viral infections. Canine parvovirus is caused by a small DNA virus and infections are often fatal in pups. Like all invertebrates, the honey bee is susceptible to many viral infections. Most viruses co-exist harmlessly in their host and cause no signs or symptoms of disease.
There are many types of plant virus, but often they cause only a loss of yield, and it is not economically viable to try to control them. Plant viruses are often spread from plant to plant by organisms, known as vectors. These are usually insects, but some fungi, nematode worms, and single-celled organisms have been shown to be vectors. When control of plant virus infections is considered economical, for perennial fruits, for example, efforts are concentrated on killing the vectors and removing alternate hosts such as weeds. Plant viruses cannot infect humans and other animals because they can reproduce only in living plant cells.
Originally from Peru, the potato has become a staple crop worldwide. The potato virus Y causes disease in potatoes and related species including tomatoes and peppers. In the 1980s, this virus acquired economical importance when it proved difficult to control in seed potato crops. Transmitted by aphids, this virus can reduce crop yields by up to 80 per cent, causing significant losses to potato yields.
Plants have elaborate and effective defence mechanisms against viruses. One of the most effective is the presence of so-called resistance (R) genes. Each R gene confers resistance to a particular virus by triggering localised areas of cell death around the infected cell, which can often be seen with the unaided eye as large spots. This stops the infection from spreading. RNA interference is also an effective defence in plants. When they are infected, plants often produce natural disinfectants that kill viruses, such as salicylic acid,nitric oxide, and reactive oxygen molecules.
Plant virus particles or virus-like particles (VLPs) have applications in both biotechnology and nanotechnology. The capsids of most plant viruses are simple and robust structures and can be produced in large quantities either by the infection of plants or by expression in a variety of heterologous systems. Plant virus particles can be modified genetically and chemically to encapsulate foreign material and can be incorporated into supramolecular structures for use in biotechnology.
Bacteriophages are a common and diverse group of viruses and are the most abundant biological entity in aquatic environments—there are up to ten times more of these viruses in the oceans than there are bacteria, reaching levels of 250,000,000 bacteriophages per millilitre of seawater. These viruses infect specific bacteria by binding to surface receptor molecules and then entering the cell. Within a short amount of time, in some cases just minutes, bacterial polymerase starts translating viral mRNA into protein. These proteins go on to become either new virions within the cell, helper proteins, which help assembly of new virions, or proteins involved in cell lysis. Viral enzymes aid in the breakdown of the cell membrane, and, in the case of the T4 phage, in just over twenty minutes after injection over three hundred phages could be released.
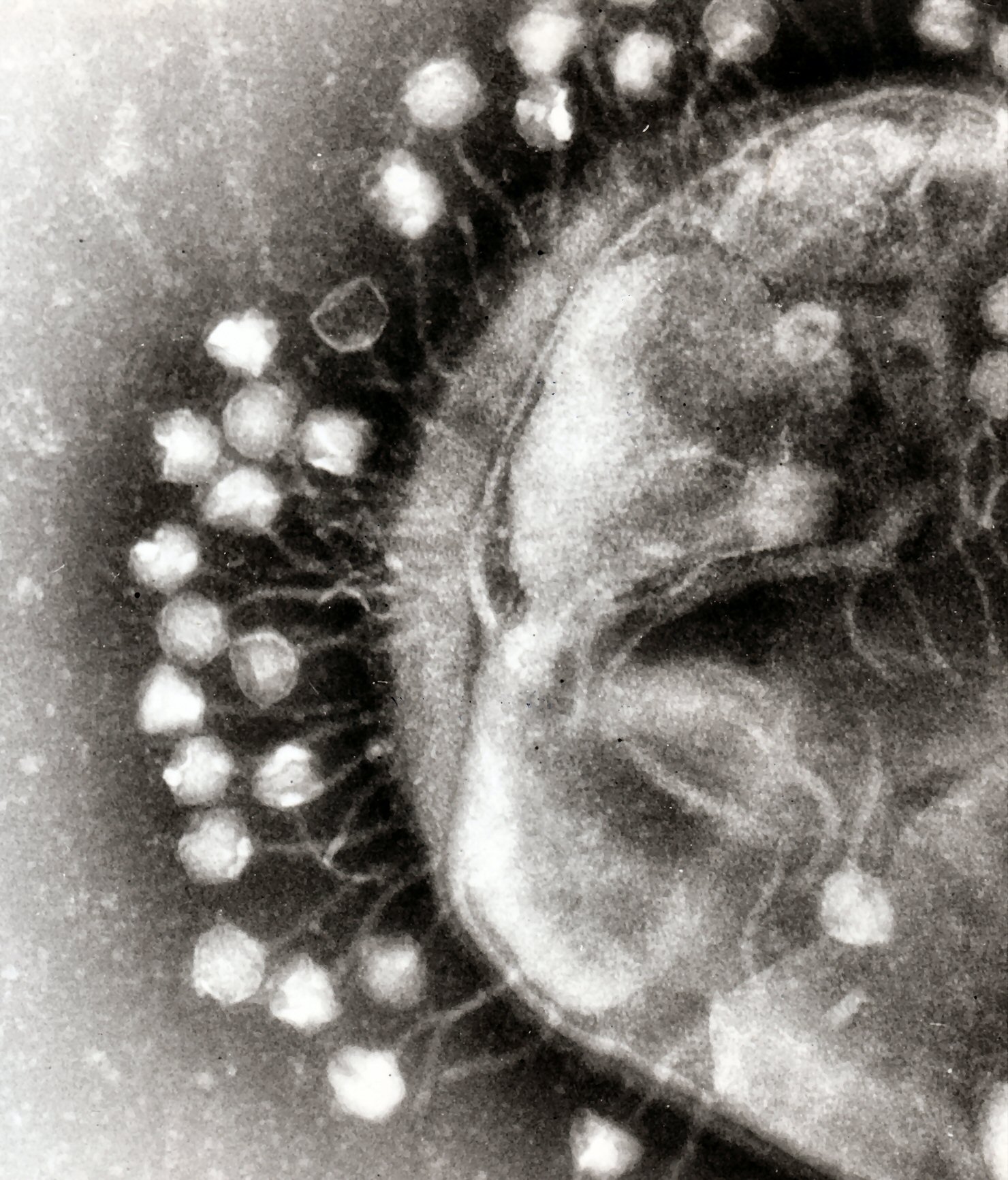
Transmission electron micrograph of multiple bacteriophages attached to a bacterial cell wal.
The major way bacteria defend themselves from bacteriophages is by producing enzymes that destroy foreign DNA. These enzymes, called restriction endonucleases, cut up the viral DNA that bacteriophages inject into bacterial cells. Bacteria also contain a system that uses CRISPR sequences to retain fragments of the genomes of viruses that the bacteria have come into contact with in the past, which allows them to block the virus's replication through a form of RNA interference. This genetic system provides bacteria with acquired immunity to infection.
Some viruses replicate within archaea: these are double-stranded DNA viruses with unusual and sometimes unique shapes. These viruses have been studied in most detail in the thermophilic archaea, particularly the orders Sulfolobales and Thermoproteales. Defences against these viruses involve RNA interference from repetitive DNA sequences within archaean genomes that are related to the genes of the viruses. Most archaea have CRISPR–Cas systems as an adaptive defence against viruses. These enable archaea to retain sections of viral DNA, which are then used to target and eliminate subsequent infections by the virus using a process similar to RNA interference.
Viruses are the most abundant biological entity in aquatic environments There are about ten million of them in a teaspoon of seawater. Most of these viruses are bacteriophages infecting heterotrophic bacteria and cyanophages infecting cyanobacteria and they are essential to the regulation of saltwater and freshwater ecosystems. Bacteriophages are harmless to plants and animals, and are essential to the regulation of marine and freshwater ecosystems are important mortality agents of phytoplankton, the base of the foodchain in aquatic environments. They infect and destroy bacteria in aquatic microbial communities, and are one of the most important mechanisms of recycling carbon and nutrient cycling in marine environments. The organic molecules released from the dead bacterial cells stimulate fresh bacterial and algal growth, in a process known as the viral shunt. In particular, lysis of bacteria by viruses has been shown to enhance nitrogen cycling and stimulate phytoplankton growth. Viral activity may also affect the biological pump, the process whereby carbon is sequestered in the deep ocean.
Microorganisms constitute more than 90% of the biomass in the sea. It is estimated that viruses kill approximately 20% of this biomass each day and that there are 10 to 15 times as many viruses in the oceans as there are bacteria and archaea. Viruses are also major agents responsible for the destruction of phytoplankton including harmful algal blooms, The number of viruses in the oceans decreases further offshore and deeper into the water, where there are fewer host organisms.
In January 2018, scientists reported that 800 million viruses, mainly of marine origin, are deposited daily from the Earth's atmosphere onto every square meter of the planet's surface, as the result of a global atmospheric stream of viruses, circulating above the weather system but below the altitude of usual airline travel, distributing viruses around the planet.
Viruses are an important natural means of transferring genes between different species, which increases genetic diversity and drives evolution. It is thought that viruses played a central role in early evolution, before the diversification of the last universal common ancestor into bacteria, archaea and eukaryotes. Viruses are still one of the largest reservoirs of unexplored genetic diversity on Earth.
Viruses are important to the study of molecular and cell biology as they provide simple systems that can be used to manipulate and investigate the functions of cells. The study and use of viruses have provided valuable information about aspects of cell biology. For example, viruses have been useful in the study of genetics and helped our understanding of the basic mechanisms of molecular genetics, such as DNA replication,transcription,RNA processing,translation,protein transport, and immunology.
Geneticists often use viruses as vectors to introduce genes into cells that they are studying. This is useful for making the cell produce a foreign substance, or to study the effect of introducing a new gene into the genome. In a similar fashion, virotherapy uses viruses as vectors to treat various diseases, as they can specifically target cells and DNA. It shows promising use in the treatment of cancer and in gene therapy. Eastern European scientists have used phage therapy as an alternative to antibiotics for some time, and interest in this approach is increasing, because of the high level of antibiotic resistance now found in some pathogenic bacteria. The expression of heterologous proteins by viruses is the basis of several manufacturing processes that are currently being used for the production of various proteins such as vaccine antigens and antibodies. Industrial processes have been recently developed using viral vectors and a number of pharmaceutical proteins are currently in pre-clinical and clinical trials.
Virotherapy involves the use of genetically modified viruses to treat diseases. Viruses have been modified by scientists to reproduce in cancer cells and destroy them but not infect healthy cells. Talimogene laherparepvec (T-VEC), for example, is a modified herpes simplex virus that has had a gene, which is required for viruses to replicate in healthy cells, deleted and replaced with a human gene (GM-CSF) that stimulates immunity. When this virus infects cancer cells, it destroys them and in doing so the presence the GM-CSF gene attracts dendritic cells from the surrounding tissues of the body. The dendritic cells process the dead cancer cells and present components of them to other cells of the immune system. Having completed successful clinical trials, the virus gained approval for the treatment of melanoma in late 2015. Viruses that have been reprogrammed to kill cancer cells are called oncolytic viruses.
Current trends in nanotechnology promise to make much more versatile use of viruses. From the viewpoint of a materials scientist, viruses can be regarded as organic nanoparticles. Their surface carries specific tools that enable them to cross the barriers of their host cells. The size and shape of viruses and the number and nature of the functional groups on their surface is precisely defined. As such, viruses are commonly used in materials science as scaffolds for covalently linked surface modifications. A particular quality of viruses is that they can be tailored by directed evolution. The powerful techniques developed by life sciences are becoming the basis of engineering approaches towards nanomaterials, opening a wide range of applications far beyond biology and medicine.
Because of their size, shape, and well-defined chemical structures, viruses have been used as templates for organising materials on the nanoscale. Recent examples include work at the Naval Research Laboratory in Washington, D.C., using Cowpea mosaic virus (CPMV) particles to amplify signals in DNA microarray based sensors. In this application, the virus particles separate the fluorescentdyes used for signalling to prevent the formation of non-fluorescent dimers that act as quenchers. Another example is the use of CPMV as a nanoscale breadboard for molecular electronics.
Many viruses can be synthesised de novo ("from scratch") and the first synthetic virus was created in 2002. Although somewhat of a misconception, it is not the actual virus that is synthesised, but rather its DNA genome (in case of a DNA virus), or a cDNA copy of its genome (in case of RNA viruses). For many virus families the naked synthetic DNA or RNA (once enzymatically converted back from the synthetic cDNA) is infectious when introduced into a cell. That is, they contain all the necessary information to produce new viruses. This technology is now being used to investigate novel vaccine strategies. The ability to synthesise viruses has far-reaching consequences, since viruses can no longer be regarded as extinct, as long as the information of their genome sequence is known and permissive cells are available. As of November 2017, the full-length genome sequences of 7454 different viruses, including smallpox, are publicly available in an online database maintained by the National Institutes of Health.
The ability of viruses to cause devastating epidemics in human societies has led to the concern that viruses could be weaponised for biological warfare. Further concern was raised by the successful recreation of the infamous 1918 influenza virus in a laboratory. Smallpox virus devastated numerous societies throughout history before its eradication. There are only two centres in the world authorised by the WHO to keep stocks of smallpox virus: the State Research Center of Virology and Biotechnology VECTOR in Russia and the Centers for Disease Control and Prevention in the United States. It may be used as a weapon, as the vaccine for smallpox sometimes had severe side-effects, it is no longer used routinely in any country. Thus, much of the modern human population has almost no established resistance to smallpox and would be vulnerable to the virus.
Introduction (B) Virus, infectious agent of small size and simple composition that can multiply only in living cells of animals,plants, or bacteria. The name is from a Latin word meaning “slimy liquid” or “poison.”
Ebolavirus.
The earliest indications of the biological nature of viruses came from studies in 1892 by the Russian scientist Dmitry I. Ivanovsky and in 1898 by the Dutch scientist Martinus W. Beijerinck. Beijerinck first surmised that the virus under study was a new kind of infectious agent, which he designated contagium vivum fluidum, meaning that it was a live, reproducing organism that differed from other organisms. Both of these investigators found that a disease of tobacco plants could be transmitted by an agent, later called tobacco mosaic virus, passing through a minute filter that would not allow the passage of bacteria. This virus and those subsequently isolated would not grow on an artificial medium and were not visible under the light microscope. In independent studies in 1915 by the British investigator Frederick W. Twort and in 1917 by the French Canadian scientist Félix H. d’Hérelle, lesions in cultures of bacteria were discovered and attributed to an agent called bacteriophage (“eater of bacteria”), now known to be viruses that specifically infect bacteria.
The unique nature of these agents meant that new methods and alternative models had to be developed to study and classify them. The study of viruses confined exclusively or largely to humans, however, posed the formidable problem of finding a susceptible animal host. In 1933 the British investigators Wilson Smith, Christopher H. Andrewes, and Patrick P. Laidlaw were able to transmit influenza to ferrets, and the influenza virus was subsequently adapted to mice. In 1941 the American scientist George K. Hirst found that influenza virus grown in tissues of the chicken embryo could be detected by its capacity to agglutinate (draw together) red blood cells.
A significant advance was made by the American scientists John Enders,Thomas Weller, and Frederick Robbins, who in 1949 developed the technique of culturingcells on glass surfaces; cells could then be infected with the viruses that cause polio (poliovirus) and other diseases. (Until this time, the poliovirus could be grown only in the brains of chimpanzees or the spinal cords of monkeys.) Culturing cells on glass surfaces opened the way for diseases caused by viruses to be identified by their effects on cells (cytopathogenic effect) and by the presence of antibodies to them in the blood. Cell culture then led to the development and production of vaccines (preparations used to elicit immunity against a disease) such as the poliovirus vaccine.
Scientists were soon able to detect the number of bacterial viruses in a culture vessel by measuring their ability to break apart (lyse) adjoining bacteria in an area of bacteria (lawn) overlaid with an inert gelatinous substance called agar—viral action that resulted in a clearing, or “plaque.” The American scientist Renato Dulbecco in 1952 applied this technique to measuring the number of animal viruses that could produce plaques in layers of adjoining animal cells overlaid with agar. In the 1940s the development of the electron microscope permitted individual virus particles to be seen for the first time, leading to the classification of viruses and giving insight into their structure.
Advancements that have been made in chemistry, physics, and molecular biology since the 1960s have revolutionized the study of viruses. For example, electrophoresis on gel substrates gave a deeper understanding of the protein and nucleic acid composition of viruses. More-sophisticated immunologic procedures, including the use of monoclonal antibodies directed to specific antigenic sites on proteins, gave a better insight into the structure and function of viral proteins. The progress made in the physics of crystals that could be studied by X-ray diffraction provided the high resolution required to discover the basic structure of minute viruses. Applications of new knowledge about cell biology and biochemistry helped to determine how viruses use their host cells for synthesizing viral nucleic acids and proteins.
📹 Lithium-oxygen storage battery; virus
Lithium-oxygen storage battery; virus
Learn how a benign bacterial virus can be used to improve the performance of lithium-oxygen storage batteries.
The revolution that took place in the field of molecular biology allowed the genetic information encoded in nucleic acids of viruses — which enables viruses to reproduce, synthesize unique proteins, and alter cellular functions — to be studied. In fact, the chemical and physical simplicity of viruses has made them an incisive experimental tool for probing the molecular events involved in certain life processes. Their potential ecological significance was realized in the early 21st century, following the discovery of giant viruses in aquatic environments in different parts of the world.
This article discusses the fundamental nature of viruses: what they are, how they cause infection, and how they may ultimately cause disease or bring about the death of their host cells. For more-detailed treatment of specific viral diseases, seeinfection.
Viruses occupy a special taxonomic position: they are not plants, animals, or prokaryotic bacteria (single-cell organisms without defined nuclei), and they are generally placed in their own kingdom. In fact, viruses should not even be considered organisms, in the strictest sense, because they are not free-living — i.e., they cannot reproduce and carry on metabolic processes without a host cell.
All true viruses contain nucleic acid — either DNA (deoxyribonucleic acid) or RNA (ribonucleic acid) — and protein. The nucleic acid encodes the genetic information unique for each virus. The infective, extracellular (outside the cell) form of a virus is called the virion. It contains at least one unique protein synthesized by specific genes in the nucleic acid of that virus. In virtually all viruses, at least one of these proteins forms a shell (called a capsid) around the nucleic acid. Certain viruses also have other proteins internal to the capsid; some of these proteins act as enzymes, often during the synthesis of viral nucleic acids. Viroids (meaning “viruslike”) are disease-causing organisms that contain only nucleic acid and have no structural proteins. Other viruslike particles called prions are composed primarily of a protein tightly complexed with a small nucleic acid molecule. Prions are very resistant to inactivation and appear to cause degenerative brain disease in mammals, including humans.
Viruses are quintessential parasites; they depend on the host cell for almost all of their life-sustaining functions. Unlike true organisms, viruses cannot synthesize proteins, because they lack ribosomes (cell organelles) for the translation of viral messenger RNA (mRNA; a complementary copy of the nucleic acid of the nucleus that associates with ribosomes and directs protein synthesis) into proteins. Viruses must use the ribosomes of their host cells to translate viral mRNA into viral proteins.
Viruses are also energy parasites; unlike cells, they cannot generate or store energy in the form of adenosine triphosphate (ATP). The virus derives energy, as well as all other metabolic functions, from the host cell. The invading virus uses the nucleotides and amino acids of the host cell to synthesize its nucleic acids and proteins, respectively. Some viruses use the lipids and sugar chains of the host cell to form their membranes and glycoproteins (proteins linked to short polymers consisting of several sugars).
The true infectious part of any virus is its nucleic acid, either DNA or RNA but never both. In many viruses, but not all, the nucleic acid alone, stripped of its capsid, can infect (transfect) cells, although considerably less efficiently than can the intact virions.
The virion capsid has three functions: (1) to protect the viral nucleic acid from digestion by certain enzymes (nucleases), (2) to furnish sites on its surface that recognize and attach (adsorb) the virion to receptors on the surface of the host cell, and, in some viruses, (3) to provide proteins that form part of a specialized component that enables the virion to penetrate through the cell surface membrane or, in special cases, to inject the infectious nucleic acid into the interior of the host cell.
Logic originally dictated that viruses be identified on the basis of the host they infect. This is justified in many cases but not in others, and the host range and distribution of viruses are only one criterion for their classification. It is still traditional to divide viruses into three categories: those that infect animals, plants, or bacteria.
Virtually all plant viruses are transmitted by insects or other organisms (vectors) that feed on plants. The hosts of animal viruses vary from protozoans (single-celled animal organisms) to humans. Many viruses infect either invertebrate animals or vertebrates, and some infect both. Certain viruses that cause serious diseases of animals and humans are carried by arthropods. These vector-borne viruses multiply in both the invertebrate vector and the vertebrate host.
Certain viruses are limited in their host range to the various orders of vertebrates. Some viruses appear to be adapted for growth only in ectothermic vertebrates (animals commonly referred to as cold-blooded, such as fishes and reptiles), possibly because they can reproduce only at low temperatures. Other viruses are limited in their host range to endothermic vertebrates (animals commonly referred to as warm-blooded, such as mammals).
The amount and arrangement of the proteins and nucleic acid of viruses determine their size and shape. The nucleic acid and proteins of each class of viruses assemble themselves into a structure called a nucleoprotein, or nucleocapsid. Some viruses have more than one layer of protein surrounding the nucleic acid; still others have a lipoprotein membrane (called an envelope), derived from the membrane of the host cell, that surrounds the nucleocapsid core. Penetrating the membrane are additional proteins that determine the specificity of the virus to host cells. The protein and nucleic acid constituents have properties unique for each class of virus; when assembled, they determine the size and shape of the virus for that specific class. The genomes of Mimiviruses and Pandoraviruses, which are some of the largest known viruses, range from 1 to 2.5 Mb (1 Mb = 1,000,000 base pairs of DNA).
Most viruses vary in diameter from 20 nanometres (nm; 0.0000008 inch) to 250–400 nm; the largest, however, measure about 500 nm in diameter and are about 700–1,000 nm in length. Only the largest and most complex viruses can be seen under the light microscope at the highest resolution. Any determination of the size of a virus also must take into account its shape, since different classes of viruses have distinctive shapes.
Shapes of viruses are predominantly of two kinds: rods, or filaments, so called because of the linear array of the nucleic acid and the protein subunits; and spheres, which are actually 20-sided (icosahedral) polygons. Most plant viruses are small and are either filaments or polygons, as are many bacterial viruses. The larger and more-complex bacteriophages, however, contain as their genetic information double-stranded DNA and combine both filamentous and polygonal shapes. The classic T4 bacteriophage is composed of a polygonal head, which contains the DNA genome and a special-function rod-shaped tail of long fibres. Structures such as these are unique to the bacteriophages.
Animal viruses exhibit extreme variation in size and shape. The smallest animal viruses belong to the families Parvoviridae and Picornaviridae and measure about 20 nm and about 30 nm in diameter, respectively. Viruses of these two families are icosahedrons and contain nucleic acids with limited genetic information. Viruses of the family Poxviridae are about 250 to 400 nm in their longest dimension, and they are neither polygons nor filaments. Poxviruses are structurally more complex than simple bacteria, despite their close resemblance. Animal viruses that have rod-shaped (helical) nucleocapsids are those enclosed in an envelope; these viruses are found in the families Paramyxoviridae,Orthomyxoviridae,Coronaviridae, and Rhabdoviridae. Not all enveloped viruses contain helical nucleocapsids, however; those of the families Herpesviridae,Retroviridae, and Togaviridae have polygonal nucleocapsids. Most enveloped viruses appear to be spherical, although the rhabdoviruses are elongated cylinders.
The criteria used for classifying viruses into families and genera are primarily based on three structural considerations: (1) the type and size of their nucleic acid, (2) the shape and size of the capsids, and (3) the presence of a lipid envelope, derived from the host cell, surrounding the viral nucleocapsid.
📹 Anatomy of virus structures
Dissect the anatomy of icosahedral, rod-shaped, and bacteriophage virus structures in electron micrographs Animation and microphotography illustrating the structural diversity of viruses.
As is true in all forms of life, the nucleic acid of each virus encodes the genetic information for the synthesis of all proteins. In almost all free-living organisms, the genetic information is in the form of double-stranded DNA arranged as a spiral lattice joined at the bases along the length of the molecule (a double helix). In viruses, however, genetic information can come in a variety of forms, including single-stranded or double-stranded DNA or RNA.
The nucleic acids of virions are arranged into genomes. All double-stranded DNA viruses consist of a single large molecule, whereas most double-stranded RNA viruses have segmented genomes, with each segment usually representing a single gene that encodes the information for synthesizing a single protein. Viruses with single-stranded genomic DNA are usually small, with limited genetic information. Some single-stranded DNA viruses are composed of two populations of virions, each consisting of complementary single-stranded DNA of polarity opposite to that of the other.
The virions of most plant viruses and many animal and bacterial viruses are composed of single-stranded RNA. In most of these viruses, the genomic RNA is termed a positive strand because the genomic RNA acts as mRNA for direct synthesis (translation) of viral protein. Several large families of animal viruses, and one that includes both plant and animal viruses (the Rhabdoviridae), however, contain genomic single-stranded RNA, termed a negative strand, which is complementary to mRNA. All of these negative-strand RNA viruses have an enzyme, called an RNA-dependent RNA polymerase (transcriptase), which must first catalyze the synthesis of complementary mRNA from the virion genomic RNA before viral protein synthesis can occur. These variations in the nucleic acids of viruses form one central criterion for classification of all viruses.
A distinctive large family of single-stranded RNA viruses is called Retroviridae; the RNA of these viruses is positive, but the viruses are equipped with an enzyme, called a reverse transcriptase, that copies the single-stranded RNA to form double-stranded DNA.
The protein capsid provides the second major criterion for the classification of viruses. The capsid surrounds the virus and is composed of a finite number of protein subunits known as capsomeres, which usually associate with, or are found close to, the virionnucleic acid.
There are two major classes of viruses based on the protein capsid: (1) those in which a single (or segmented) linear nucleic acid molecule with two free ends is essentially completely extended or somewhat coiled (a helix) and (2) those in which the nucleic acid, which may or may not be a covalently closed circle, is wound tightly into a condensed configuration, like a ball of yarn. These two classes of virus assume in the first case a long, extended rodlike structure and in the second case a symmetrical polygon.
By far the best-studied example of a helical rod-shaped virus is the tobacco mosaic virus, which was crystallized by Wendell Stanley in 1935. The tobacco mosaic virus contains a genome of single-stranded RNA encased by 2,130 molecules of a single protein; there are 16(1/3) protein molecules for each turn of the RNA helix in the ratio of three nucleotides for each protein molecule.
Tobacco mosaic virus
Schematic structure of the tobacco mosaic virus. The cutaway section shows the helical ribonucleic acid associated with protein molecules in a ratio of three nucleotides per protein molecule.
Under the right environmental conditions, viral RNA and protein molecules in liquid suspension will assemble themselves into a perfectly formed and fully infectious virus. The length of the helical virus capsid is determined by the length of the nucleic acid molecule, which is the framework for the assembly of the capsid protein. The various helical viruses have different lengths and widths, depending on the size of the nucleic acid as well as on the mass and shape of the protein molecule. Some of these helical viruses form rigid rods, while others form flexible rods, depending on the properties of the assembled proteins.
Polygonal viruses vary greatly in size, from 20 to 150 nm in diameter, essentially proportional to the size of the nucleic acid molecule coiled up inside the virion. Most, if not all, of the polygonal viruses are icosahedral; like a geodesic dome, they are formed by equilateral triangles, in this case 20. Each triangle is composed of protein subunits (capsomeres), often in the form of hexons (multiples of six) that are the building blocks of the capsid. There are 12 vertices (the apical junctions of these 20 triangles), each comprising a penton (five subunits). These icosahedral virions have three axes of fivefold, threefold, and twofold rotational symmetry. The number of capsomeres is a basis for taxonomic classification of these virus families. Certain icosahedral viruses, usually those that are more complex, contain internal proteins adhering to the nucleic acid that are not accessible at the surface of the virions.
Virus icosahedron
A virus icosahedron (20-sided structure) shown in the (left) twofold, (centre) threefold, and (right) fivefold axes of symmetry. Edges of the upper and lower surfaces are drawn in solid and broken lines, respectively..
Surrounding viruses of either helical or icosahedral symmetry are lipoprotein envelopes, unit membranes of two lipid layers interspersed with protein molecules (lipoprotein bilayer). These viral membranes are composed of phospholipids and neutral lipids (largely cholesterol) derived from cell membranes during the process known as budding. Virtually all proteins of the cell membrane, however, are replaced by proteins of viral origin during budding. Although all the viral envelope lipids originate from the cell, their relative proportions vary from those in the cell membrane because the viral proteins select only certain lipids during budding.
Associated with the virion membrane are “integral” glycoproteins, which completely traverse the lipid bilayer, and “peripheral” matrix proteins, which line the inner surface. The glycoproteins contain regions of amino acids that, in the first step of viral infection, recognize host-cell receptors. Matrix proteins appear to function in the selection of regions of the cell membrane to be used for the viral membrane, as well as to aid the virion in entering cells.
The Cycle Of Infection
The Cycle Of Infection
The Cycle Of Infection (B)
Viruses can reproduce only within a host cell. The parental virus (virion) gives rise to numerous progeny, usually genetically and structurally identical to the parent virus. The actions of the virus depend both on its destructive tendencies toward a specific host cell and on environmental conditions. In the vegetative cycle of viral infection, multiplication of progeny viruses can be rapid. This cycle of infection often results in the death of the cell and the release of many virus progeny. Certain viruses, particularly bacteriophages, are called temperate (or latent) because the infection does not immediately result in cell death. The viral genetic material remains dormant or is actually integrated into the genome of the host cell. Cells infected with temperate viruses are called lysogenic because the cells tend to be broken down when they encounter some chemical or physical factor, such as ultraviolet light. In addition, many animal and plant viruses, the genetic information of which is not integrated into the host DNA, may lie dormant in tissues for long periods of time without causing much, if any, tissue damage. Viral infection does not always result in cell death or tissue injury; in fact, most viruses lie dormant in tissue without ever causing pathological effects, or they do so only under other, often environmental, provocations.
📹 Anatomy of virus structures
Study how bacteriophages replicate by injecting nucleic acid into a bacteria cell to create virionsThe cycle of infection results in the death of the host cell and the release of many virus particles, called virions.
Although the reproductive pathways of different viruses vary considerably, there are certain basic principles and a particular series of events in the cycle of infection for most, if not all, viruses. The first step in the cycle of infection is that the invading parental virus (virion) must attach to the surface of the host cell (adsorption). In the second step, the intact virion either penetrates the outer membrane and enters the cell’s interior (cytoplasm) or injects the genetic material of the virus into the interior of the cell while the protein capsid (and envelope, if present) remains at the cell surface. In the case of whole-virion penetration, a subsequent process (uncoating) liberates the genetic material from the capsid and envelope, if present. In either case, the viral genetic material cannot begin to synthesize protein until it has emerged from the capsid or envelope.
Virus: invasion of a cell The process by which a virus invades a cell and reproduces..
Certain bacterial viruses, such as the T4 bacteriophage, have evolved an elaborate process of infection: following adsorption and firm attachment of the virus’s tail to the bacterium surface by means of proteinaceous “pins,” the musclelike tail contracts, and the tail plug penetrates the cell wall and underlying membrane and injects virus (phage) DNA into the cell. Other bacteriophages penetrate the cell membrane by different means, such as injecting the nucleic acid through the male (sex) pili of the bacterium. In all bacterial viruses, penetration transmits the viral nucleic acid through a rigid bacterial cell wall
Bacteriophage General structure of T4 bacteriophage and a model of its mode of attachment to, and injection of its DNA into, a bacterial cell.
Plant cells also have rigid cell walls, which plant viruses cannot ordinarily penetrate. Plant viruses, however, have not evolved their own systems for injecting nucleic acids into host cells, and so they are transmitted by the proboscis of insects that feed on plants. In the laboratory, plant viruses penetrate plant cells if the cell walls have been abraded with sandpaper or if cell protoplasts (plasma membrane, cytoplasm, and nucleus) are devoid of walls.
Penetration of animal cells by viruses involves different processes, because animal cells are enclosed not by walls but by a flexible lipoprotein bilayer membrane. Most animal viruses, whether or not they are encased in lipid envelopes, penetrate cells in an intact form by a process called endocytosis. The membrane invaginates and engulfs a virus particle adsorbed to a cell, usually in an area of the membrane called a coated pit, which is lined by a special protein known as clathrin. As the coated pit invaginates, it is pinched off in the cytoplasm to form a coated vesicle. The coated vesicle fuses with cytoplasmic endosomes (membrane-enclosed vesicles) and then with cell organelles called lysosomes, which are membrane-enclosed vesicles containing enzymes. In an acidic environment, the membrane of an enveloped virus fuses with the endosome membrane, and the viral nucleocapsid is released into the cytoplasm. Nonenveloped viruses presumably undergo a similar process, by which the protein capsid is degraded, releasing the naked viral nucleic acid into the cytoplasm.
Endocytosis
Adsorption to and entry into a cell of an enveloped animal virus by the process of endocytosis into clathrin-coated vesicles, which fuse with large vesicles (endosomes and lysosomes). The process triggered by the viral glycoprotein results in fusion and release of the viral nucleocapsid into the cytoplasm.
The order of the stages of viral replication that follow the uncoating of the genome varies for different virus classes. For many virus families the third step in the cycle of infection is transcription of the genome of the virus to produce viral mRNA, followed by the fourth step, translation of viral mRNA into proteins. For those viruses in which the genomic nucleic acid is an RNA that can serve as a messenger (i.e., positive-strand RNA viruses), the third step is the translation of the RNA to form viral proteins; some of these newly synthesized viral proteins are enzymes that synthesize nucleic acids (polymerases), which carry out a fourth step, the transcription of more mRNA from the viral genome. For the more complicated DNA viruses, such as adenoviruses and herpesviruses, some regions of the genome synthesize “early” mRNAs, which are translated into polymerases that initiate the transcription of “late” regions of the DNA into mRNAs, which are then translated into structural proteins.
Regardless of how the third and fourth steps proceed, the fifth step in the cycle of infection is replication (reproduction of the parental genome to make progeny genomes). The sixth step is the assembly of the newly replicated progeny genomes with structural proteins to make fully formed progeny virions. The seventh and last step is the release of progeny virions by lysis of the host cell through the process of either extrusion or budding, depending on the nature of the virus. In a host animal or cell culture, this seven-step process may be repeated many times; the progeny virions released from the original site of infection are then transmitted to other sites or to other individuals.
For most animal and plant RNA viruses, all replicative events take place in the cytoplasm; in fact, many of these RNA viruses can grow in host cells in which the nucleus has been removed. Replication of most animal and plant DNA viruses, as well as the RNA influenza virus, takes place in the nucleus. In these viruses, transcription takes place in the nucleus, the mRNA migrates to the cytoplasm, where it is translated, and these viral proteins migrate back to the nucleus, where they assemble with newly replicated progeny genomes. Migration of newly translated viral proteins from the cytoplasm to the nucleus is generally a function of specific amino acid sequences called “signals,” which translocate the protein through pores in the nucleus membrane.
Many bacterial and animal viruses lie dormant in the infected cell, and their DNA may be integrated into the DNA of the host cell chromosome. The integrated viral DNA replicates as the cell genome replicates; after cell division, the integrated viral DNA is duplicated and usually distributed equally to the two cells that result. The bacteria that carry the noninfective precursor phage, called the prophage, remain healthy and continue to grow until they are stimulated by some perturbing factor, such as ultraviolet light. The prophage DNA is then excised from the bacterial chromosome, and the phage replicates, producing many progeny phages and lysing the host bacterial cell. This process, originally discovered in temperate bacteriophages in 1950 by the French microbiologist André Lwoff, is called lysogeny.
The classic example of a temperate bacteriophage is called lambda (λ) virus, which readily causes lysogeny in certain species of the bacterium Escherichia coli. The DNA of the λ bacteriophage is integrated into the DNA of the E. coli host chromosome at specific regions called attachment sites. The integrated prophage is the inherited, noninfectious form of the virus; it contains a gene that represses the lytic functions of the phage and thus ensures that the host cell will continue to replicate the phage DNA along with its own and that it will not be destroyed by the virus. Ultraviolet light, or other factors that stimulate the replication of DNA in the host cell, causes the formation of a recA protease, an enzyme that breaks apart the λ phage repressor and induces λ phage replication and, eventually, destruction of the host cell.
Excision of the prophage DNA from the host chromosomal DNA (as an initial step in the synthesis of an infective, lytic virus) sometimes results in the removal of some of the host cell DNA, which is packaged into defective bacteriophages; part of the bacteriophage DNA is removed and replaced at the other end by a gene of the host bacterium. Such a virus particle is called a transducing phage because, when it infects a bacterial cell, it can transmit the gene captured by λ phage DNA into the next bacterial cell it infects. Transduction by bacteriophages is an efficient means for transferring the genetic information of one bacterial cell to another.
This means of transferring genetic information, called lysogenic conversion, imparts genes with special functions to bacterial cells without such functions. It is common in bacteria and is an important aspect of the epidemiology (incidence, distribution, and control) of infectious diseases. For example, the bacterium Corynebacterium diphtheriae is the causative agent of diphtheria, but only when it contains the prophage of bacteriophage β, which codes for the toxin that is responsible for the disease.
A phenomenonanalogous to bacterial celllysogeny occurs in animal cells infected with certain viruses. These animal viruses do not generally cause disease immediately for certain animal cells. Instead, animal cells are persistently infected with such viruses, the DNA of which (provirus) is integrated into the chromosomal DNA of the host cell. In general, cells with integrated proviral DNA are converted into cancer cells, a phenomenon known as malignant transformation. As is the case with bacterial prophages, the transformed animal cell contains no infectious virus but only the integrated provirus DNA, which replicates along with the dividing cell’s chromosomes. Therefore, following mitosis of the transformed cell, each new cell receives a copy of the proviral DNA. The hallmark of these transformed animal cells is that their growth is uncontrollable; unlike normal cells, their growth is not inhibited by contact with other cells, and they lose their capacity to adhere (anchor) to certain surfaces. Growth of normal tissues and organs is also controlled by a genetic phenomenon called programmed cell death, or apoptosis, in which a certain number of cells will die and be eliminated after a finite number of divisions. Malignant transformation can impede programmed cell death, thus allowing the cells to grow uncontrolled and resulting in cancer.
Among the animal viruses that cause malignant transformation by integration of proviral DNA are several families of DNA viruses and one large family of RNA viruses, the Retroviridae. Viruses of the family Polyomaviridae, a group of papovaviruses, were perhaps the first to be associated with malignancy (causing death or illness) in animals. Polyomaviruses are widespread in mice; they can infect other rodents, and they can cause tumours in infected animals. Another virus of the family Polyomaviridae is simian virus 40 (SV40), originally isolated from cells of the African green monkey (Cercopithecus sabaeus), where it grows rapidly and kills the cells. Infection of rodent or human cells, however, results in an abortive infection (an incompatibility between the virus and the host cell) but sometimes induces malignancy (sarcomas or lymphomas) in the occasional cell that is transformed. Viruses related to polyomavirus and SV40 have been isolated from humans, one of which, the JC virus, appears to be the causative agent of a fatal neurological disease called progressive multifocal leukoencephalopathy. In general, however, the human papovaviruses are not clearly associated with disease.
Other papovaviruses include the papillomaviruses (family Papillomaviridae), which are also small polygonal viruses containing circular double-stranded DNA. The papillomaviruses are associated with usually benign (nonthreatening) but widespread tumours, called papillomas or polyps, occurring in human skin and the genital tract. Specific papillomaviruses have been identified in humans in common warts and in genital warts (condylomata acuminata). Cancers of the human genital tract, particularly uterine cancer of the cervix, are frequently found in association with human papillomavirus type 16 (HPV 16); the virus undoubtedly is transmitted as a venereal disease.
Certain viruses of the family Adenoviridae, originally found in the tonsils and adenoids of humans, cause malignant transformation in certain cells. This phenomenon of cancer induction under laboratory conditions has been studied widely, but there is no evidence that the common adenoviruses cause cancers in humans. The common viruses of the family Herpesviridae, however, including the common herpes simplex viruses that cause cold sores and the venereal disease genital herpes, are suspected of being causative agents of cancer. Like the adenoviruses, the herpesviruses can cause malignant transformations, and their DNA is integrated into the host cell chromosome. A herpesvirus known as the Epstein-Barr virus causes a frequently fatal childhood cancer called Burkitt lymphoma as well as the nonmalignant disease infectious mononucleosis. The herpesvirus cytomegalovirus lies dormant in the tissues of most humans and can be induced to cause fatal diseases in infants and immunocompromised adults. A different herpesvirus causes chickenpox (varicella); the same virus lies latent in the tissues for long periods of time (perhaps years or decades) and later undergoes recrudescence (the recurrence of symptoms after they have abated) to cause the painful skin and neurological disease called herpes zoster, or shingles. In addition, there are herpesviruses that cause disease in animals—for example, the widespread and usually fatal disease in chickens called Marek’s disease. The widespread distribution of viruses of the family Herpesviridae is evident from other diseases in monkeys and frogs.
Adenovirus
The structure of an adenovirus showing its components and the polypeptides (proteins) exhibited by staining after electrophoresis.
The viruses of the family Retroviridae are perhaps the most widely distributed of the transforming viruses that infect eukaryotic cells ranging from yeast to humans. It was suggested early in the 20th century that viruses cause leukemias and lymphomas in birds. In 1911 the American pathologist Peyton Rous first described a virus that causes sarcomas in chickens.
The virions of retroviruses are spherical (or polygonal) and are surrounded by a lipid membrane containing a glycoprotein that recognizes and binds to cell receptors of a particular species (type-specific glycoproteins). Retrovirus genomes consist of two identical RNA molecules, each with 7,000 to 10,000 nucleotides. Associated with the virion RNA is an enzyme, an RNA-dependent DNA polymerase, also called a reverse transcriptase. Using the virion RNA as a template, the reverse transcriptase catalyzes the synthesis of a linear DNA molecule complementary to the virion RNA. The new complementary strand of DNA also serves as a template for the reverse transcriptase, which makes a second anticomplementary DNA molecule, thus forming double-stranded DNA. The genomic RNA of fully infectious bird retroviruses, those that can replicate autonomously, has four genes that code sequentially for group-specific antigens, the reverse transcriptase, the envelope glycoprotein, and the sarcoma-transforming protein. At each end of the genome are homologous flanking nucleotide sequences, known as long terminal repeats (LTR), which code for double-stranded DNA that can recognize host cell DNA sequences for integration of the proviral DNA into the host cell chromosome. Many retroviruses are defective and cannot replicate in cells without helper (nondefective) retroviruses. The helper retroviruses generally transform fibroblastic cells, resulting in malignant sarcomas, whereas the defective retroviruses transform blood-cell precursors, resulting in leukemias.
Many different retroviruses have been identified as causative agents of cancers in birds, rodents (particularly mice), domestic cats, monkeys, and humans. Certain lymphatic leukemias in humans are caused by human T-cell leukemia virus (HTLV); acquired immune deficiency syndrome (AIDS) is caused by a retrovirus called human immunodeficiency virus (HIV).
Retroviruses originated from genes in many different species of animals and even lower forms of life. Individual retroviruses are limited in their host range and do not readily cross species barriers. Virtually every retrovirus studied to date is analogous to the genes normally found in animals (including humans), known as proto-oncogenes, genes that are involved with regulating normal cell growth and development and that also have the potential to change into cancer-causing genes. These proto-oncogenes have deoxynucleotide sequences closely, but not entirely, homologous (i.e., of the same type and order) to the nucleotide sequences of a corresponding viral cancer-causing gene, called an oncogene. Integration of retrovirus DNA into cell chromosomes results in cancer, but the proto-oncogenes do not become cancer-causing genes unless triggered by another event. Cancers caused by chemical or physical carcinogens in the environment probably often, if not invariably, are due to alterations in the sequences of proto-oncogenes that have converted them to oncogenes. Some of the DNA tumour viruses, such as SV40 or adenoviruses, may induce malignant transformation when their DNA is integrated in proximity to the site of a proto-oncogene. All cancers studied to date appear to be due to either mutations in proto-oncogenes or the inheritance of mutated tumour suppressor genes, which normally regulate the function of proto-oncogenes.
Although viruses were originally discovered and characterized on the basis of the diseases they cause, most viruses that infect bacteria, plants, and animals (including humans) do not cause disease. In fact, bacteriophages may be helpful in that they rapidly transfer genetic information from one bacterium to another, and viruses of plants and animals may convey genetic information between similar species, helping their hosts survive in hostile environments. In the future this could also be true for humans. Recombinant DNA biotechnology shows great promise for the repair of genetic defects. Afflicted persons are injected with cells transformed by viruses that carry a functional copy of the defective human gene. The virus integrates the normal gene into the DNA of the human cell.
Of those viruses that cause disease, some cause short-term (acute) diseases and others recurring or long-term (chronic) diseases. Some viruses cause acute disease from which there is fairly rapid recovery but may persist in the tissues, remaining dormant for long periods of time, and then become active again, bringing about serious disease decades later. Slowly progressive viruses have long incubation periods before the onset of disease. As mentioned above, the DNA of certain viruses becomes integrated into the genome of the host cell, often resulting in malignant transformation of cells, which become cancers.
The nature of the disease caused by a virus is generally a genetic property of the virus as well as of the host cells. Many viruses, however, can remain dormant in the tissues of the host (latency). Viruses that cause acute disease are generally, but not always, those that rapidly harm or destroy cells (cytopathic effects) and have the capacity to shut off protein or nucleic acid synthesis within the host cell.
Human poliovirus and related picornaviruses that infect other animal species are examples of acute infectious agents that shut down protein synthesis in the host cell soon after infection; these picornaviruses also inhibit cellular RNA and DNA synthesis. Another virus that rapidly kills the infected cell is the negative-strand vesicular stomatitis virus (VSV) of the family Rhabdoviridae; viral RNA newly synthesized by infectious VSV rapidly shuts off cellular RNA synthesis and, to a somewhat lesser extent, cellular protein synthesis. In both poliovirus and VSV, the infected cell dies within hours of the inhibition of cellular RNA and protein synthesis. Influenza A viruses of the family Orthomyxoviridae, which cause a highly contagiousrespiratory disease in humans, inhibit cellular macromolecular synthesis by several unique mechanisms, including blocking the maturation of cellular mRNAs and cleaving off the ends of cellular mRNAs in the nucleus of infected cells. Other viruses that inhibit cellular macromolecule synthesis and produce acute infections include the poxviruses,reoviruses,togaviruses,adenoviruses, and herpesviruses; the latter two persist in host tissues for long periods of time and cause chronic infection as well.
Many, if not most, diseases resulting from viral infection of vertebrates are caused not by a direct effect of the virus but rather by a secondary immune response. Essentially all viral proteins are recognized by vertebrate animals as immunologically foreign, and the immune systems of these animals mount two kinds of immune response, humoral and cellular. In humoral immunity,B lymphocytes, usually triggered by helper T lymphocytes, make antibodies (proteins that recognize and bind foreign molecules) to the viral protein. The antibody synthesized as a result of the immune response against a specific viral antigen usually benefits the infected host because that antibody can neutralize the infectivity of the specific virus in the blood and tissues of the infected host. Viruses inside the cell are not accessible to the antibody, because it cannot cross the cell membrane barrier.
In cellular immunity, a killer T cell recognizes and kills a virus-infected cell because of the viral antigen on its surface, thus aborting the infection because a virus will not grow within a dead cell. If the virus-infected cells are not essential for host functions, the killer T cell can prevent the spread of the infecting virus to other cells and distant tissues. Not infrequently, the virus-specific T lymphocyte kills vital cells such as nerve cells (neurons),muscle cells, and liver cells, all of which carry out important functions. In addition, the death of cells results in an inflammatory response, which also can damage vital tissues. Therefore, the cellular immune response to a viral infection can cause disease. In general, diseases caused by chronic viral infections, but also occasionally by subacute (between acute and chronic) viral infections, are caused by cellular immune responses that damage the virus-infected tissue.
Cytotoxic T cell
A cytotoxic T cell (left) recognizes antigens on the surface of a cell infected with a virus (right), enabling the T cell to bind to and kill the infected cell.
Acute viral infections are of two types—local and systemic—both usually resulting from a direct effect of the invading virus on host tissue cells. Acute local infections generally occur at the site of viral infection. For example, acute respiratory infections include (1) the common cold, in which the rhinovirus infects only the nasal mucosa, (2) influenza, in which the virus is found in both nasal and bronchial mucosa, where severe damage can result in death, (3) flulike illnesses caused by adenoviruses localized in lymphoid tissue of the throat (although infection also can occur in the intestine and the eye or be spread to the heart), and (4) severe respiratory infections of infants and children, caused by parainfluenza viruses or respiratory syncytial viruses, which may be life-threatening. Examples of acute infections localized to the intestine include those that result in enteritis (bowel inflammation), which may be accompanied by diarrhea; these are often caused by rotaviruses and coronaviruses.
Many viruses transmitted by the respiratory route (from sneezes and coughs, for example) and limited to humans begin their cycle of infection in the upper respiratory tract (nose and throat) and then enter the bloodstream, where they are spread to distant tissues. Examples of such diseases are measles,mumps, and chickenpox, in which the growth of the specific virus in the mucosal cells of the throat during the first few days of infection usually results in mild fever and achiness; this stage is called the prodromal period of the illness. During the next few days, the virus enters the draining lymph nodes and then the bloodstream, where it is spread throughout the tissues of the body, resulting in fever and rash (in the case of measles and chickenpox) and inflammation of the parotid glands and, less frequently, the testes, ovaries, and joints (in the case of mumps). Varicella (chickenpox) virus rarely causes pneumonia, but all these viruses can cause meningitis and, rarely, encephalitis. A similar pattern of infection formerly occurred with smallpox, a disease that was more frequently fatal but now ostensibly has been eradicated.
A large number of viruses of the digestive tract (enteroviruses)—among them poliovirus, Coxsackie viruses, and echoviruses (enteric cytopathic human orphan virus)—also cause a two-phase illness. Enteroviruses grow initially in the intestinal tract and are transmitted by mouth through water, food, and other materials contaminated with feces. The viruses are resistant to the acid normally found in the stomach and thus reach the intestinal tract, where they multiply in living mucosal cells. This initial period of viral invasion and growth in the intestine causes either an initial mild febrile illness or is asymptomatic. Over the next few days these enteroviruses are spread from the intestinal mucosa to the draining lymph nodes, from which they invade the bloodstream, resulting in a condition known as viremia. From the bloodstream the viruses are widely spread to all tissues, but in most cases no symptomatic disease occurs. Poliovirus in less than 1 percent of cases affects the spinal cord or brain, resulting in paralysis or death. Different types of Coxsackie viruses and echoviruses can cause acute, usually nonfatal, illnesses such as meningitis, carditis, pleurisy, or rashes. Enteroviruses have also been linked to acute flaccid myelitis, a polio-like disease characterized by sudden muscle weakness and paralysis.
Many viral diseases are transmitted by bites of insects or other arthropods, and these infections usually begin in the skin or lymph nodes and rapidly invade the bloodstream. The nature of the disease caused by these arthropod-borne viruses (arboviruses) is determined by the affinity (tropism) of each virus for specific organs. Many that have an affinity for brain tissue cause encephalitis or meningitis, but others primarily infect the muscles, liver, heart, or kidneys. Virtually all these diseases are epidemic in character, and the viruses that cause them are the primary pathogens of birds and mammals. The insect, usually a certain species of mosquito, takes a blood meal from the infected host bird or mammal and shortly thereafter bites a human, thus transmitting the virus. These arboviruses do not ordinarily multiply in the insect but simply reside on its proboscis. Examples of human epidemic diseases resulting from transmission of these often fatal arboviruses are encephalitis caused by viruses of the family Togaviridae and Flaviviridae,yellow fever and dengue caused by viruses of the family Flaviviridae, and hemorrhagic fevers caused by viruses of the families Bunyaviridae and Arenaviridae. Of considerable interest and concern is the identification of new strains of viruses, particularly a hantavirus of the Bunyaviridae family that was responsible for an epidemic in the early 1990s in the southwestern United States that resulted in considerable numbers of fatal human infections.
Inapparent infections (those that do not cause specific signs and symptoms) often result after exposure to picornaviruses,influenza viruses, rhinoviruses,herpesviruses, and adenoviruses but less frequently to measles and chickenpox viruses. In cases of inapparent infection, long-lasting immunity develops, but only to the strain of virus that has the same antigenic composition as the original infecting virus.
Certain of these viruses persist in the tissues of the host after the initial infection despite the presence of circulating antibodies to it in the blood and tissues. Such viruses probably reside inside cells, where they are protected from antibodies that cannot penetrate the cell membrane. Among persistent viruses are adenoviruses, measles virus, and, in particular, many kinds of herpesviruses. The genetic information of herpesviruses and adenoviruses can be integrated into the genome of the host cell, but it is believed that these viruses frequently, and the measles virus invariably, reside in cells in the form of extrachromosomal genes (genes not integrated in chromosomes). These dormant viruses can be activated by many factors, such as trauma, another infection, emotional stress, menstruation, excessive exposure to sunlight, and various illnesses.
The phenomenon of latency and reactivation is particularly common among viruses of the family Herpesviridae, which cause chronic or recurrent diseases: (1) herpes simplex virus type 1, which causes recurrent cold sores, (2) herpes simplex virus type 2 in genital tissue, which causes repeated herpetic infections of the vagina or penis, (3) cytomegalovirus, which usually produces an inapparent infection activated by simultaneously occurring disease to cause severe liver, lung, or nervous-system disease, and (4) varicella virus, which is the causative agent of chickenpox but which can be activated decades later to produce herpes zoster (shingles). A rare, but invariably fatal, disease of the nervous system is subacute sclerosing panencephalitis (SSPE), which is a progressive degenerative condition caused by measles virus (a paramyxovirus) lying dormant in brain cells for many years and then reactivated, usually in adolescence. There is no simple explanation for why latent viruses, such as those in the family Herpesviridae, that are present in the tissues of most adult humans can be activated to cause disease in some people but not in others.
Although some viruses multiply slowly, this is not always the explanation for the chronicity or the slow progression of the diseases caused by these viruses. Hepatitis, for example, is a subacute or chronic disease, with a long latent period, that is caused by at least five viruses with different properties. HepatitisA is caused by a picornavirus usually transmitted by the fecal-oral route in a manner similar to that of poliovirus. Hepatitis B is caused by a small DNA virus that contains its own DNA polymerase and is transmitted by transfusion of blood and other blood products, by the sharing of nonsterile hypodermic needles among drug users, by sexual intercourse, or from mother to neonate. Hepatitis B virus is classified with similar viruses of birds in the family Hepadnaviridae. Most cases of hepatitis spread by the transfusion of blood or blood products or by needles shared by drug users are caused by a third, completely distinct virus—originally called non-A, non-B hepatitis but now known to be a member of the virus family Flaviviridae—designated hepatitis C virus. A fourth unique agent that causes hepatitis is designated hepatitis delta virus, which has not yet been classified taxonomically but is a small enveloped virus containing a circular RNA genome; hepatitis B virus serves as a helper for replication of hepatitis delta virus, the virions of which contain hepatitis B surface antigen (HBsAg). The fifth causative agent of viral hepatitis, largely occurring in Asia and Africa, is a small RNA virus tentatively classified as a member of the family Caliciviridae and designated hepatitis E virus.
Many other agents that appear to cause chronic and slowly progressive diseases, particularly those affecting the nervous system, have been identified. A fatal neurological disorder of sheep, called scrapie, has an incubation period of years and may be caused by a heat-resistant protein called a prion, which is self-replicating. Similar, rather obscure agents have been identified for two uncommon fatal disorders of the nervous system called Creutzfeldt-Jakob disease and kuru.
The disease now known as AIDS was first recognized in homosexuals and hemophiliacs about 1981 and continues to be disseminated throughout the world to become one of the most devastating epidemics of all time. AIDS is caused by HIV, a member of a genetically more complex group of the family Retroviridae called lentiviruses. Closely related viruses of monkeys and cats cause similar diseases. HIV is transmitted by blood and other body fluids and infects primarily helper T lymphocytes and other cells with CD4 surface receptors (cell surface proteins that react with antigens), to which the virus binds. After the virus has been dormant for years, destruction of T lymphocytes results in drastic depression of the immune system. Death almost invariably results from “opportunistic” infections such as pneumonia—caused by ordinarily nonpathogenic organisms such as Pneumocystis carinii—or tuberculosis or by cancers such as Kaposi sarcoma and lymphomas.
The spread of many viral diseases can be prevented by hygienic factors such as efficient sanitation facilities, effective waste disposal, clean water, and personal cleanliness. Active immunization by vaccines (antigen-containing preparations that elicit the synthesis of antibodies and thus immunity) has been useful in preventing common epidemics caused by acutely infectious viruses.
The best example of such a preventable disease is smallpox, caused by a disease-producing virus that at one time was found worldwide. In 1796 the English physician Edward Jenner discovered that the milder cowpox virus could serve as a live vaccine (an antigenic preparation consisting of viruses whose disease-producing capacity has been weakened) for preventing smallpox; Jenner published his findings in 1798. The program of vaccination that resulted from Jenner’s discovery is one of the greatest success stories in the annals of medicine; in 1980 the World Health Organization declared that the disease had been eliminated.
A different prospect is presented by rabies, an invariably fatal viral disease mentioned in ancient Greek literature. Transmitted by the bite of dogs and other domestic and wild animals, the rabies virus is more difficult to eradicate because it is present in wild animals throughout the world, except in certain island countries such as Great Britain and Australia. Influenza virus is also distributed worldwide, but, of the three major immunologic types, only one (type A) is responsible for large epidemics. The worldwide epidemic (pandemic) of influenza at the end of World War I is estimated to have caused 20 million deaths, mostly of adolescents and young adults. Because of virus mutations that produce minor antigenic changes every year and major antigenic shifts about every 10 years, influenza viruses have the capacity to resist inactivation by antibodies acquired by previous infection or vaccination. Development of effective vaccines to combat influenza is a difficult task, although existing vaccines are partially effective and are recommended for people at high risk—i.e., the elderly and those with chronic disease of the respiratory or circulatory systems.
Vaccines are most successful when directed against those viruses that do not mutate and that infect only humans. In addition to smallpox, a successful vaccine program has been carried out against polio. Polioviruses exist in only three antigenic types, each of which has not changed significantly for decades. The vaccines available are the “killed” (Salk) vaccine, composed of inactivated virus of the three types, and the “live” (Sabin) vaccine, composed of genetically attenuated viruses of the three types. In developed countries these vaccines, which were introduced in the 1950s, have lowered the incidence of paralysis resulting from polio. The disease still occurs in developing countries and recurs in some developed countries where vaccination programs have not been enforced. Rare cases of polio occur from the Sabin vaccine strain of type-3 poliovirus, which is genetically unstable and occasionally reverts to the virulent form.
Vaccination can prevent diseases caused by strictly human viruses that exist in only one antigenic and stable type. Measles has been prevented in developed countries with routine vaccination. Measles, however, may still be the major cause of death in children in developing countries. Vaccination for mumps and chickenpox promises to be successful because the causative viruses of these diseases show little tendency to vary antigenically and are confined to humans. On the other hand, development of vaccines for the common cold caused by rhinoviruses, similar to polioviruses, will be a formidable, if not impossible, task because there are at least 100 antigenic types of the rhinovirus. Rhinoviruses and paralysis-inducing enteroviruses, however, depend on a protein in host cells called methyltransferase SETD3 for their replication; this discovery raised the possibility of someday being able to suppress the protein therapeutically to protect individuals against infection by these viruses.
Also daunting is the task of developing a vaccine against HIV. The major antigenic component of this virus is a surface-membrane-inserted glycoprotein (gp120), which has a startling rate of mutation. The extreme antigenic diversity that results from the mutability of the gene that codes for this protein would prevent HIV from being identified and attacked by circulating antibodies or killer T lymphocytes.
Unlike bacteria, viruses mimic the metabolic functions of their host cells. Antibiotics are not effective against viruses. It is difficult to identify chemical compounds that inhibit the multiplication of viruses but do not slow the functions of, or are not toxic to, the host cell. Despite this difficulty, an effective antiviral drug has been developed against influenza virus. This drug targets a viral enzyme called the neuraminidase and is orders of magnitude less active against nonviral neuraminidases. These neuraminidase inhibitors are most effective when administered prophylactically or within the first 30 hours of symptom onset and can be used to limit the spread of influenza virus and to complement the administration of vaccines. Other chemicals that exert a selectively greater effect on viral replication than they do on cell replication include ribavirin, acyclovir, and zidovudine (azidothymidine [AZT]). These drugs have been partially effective in improving, if not curing, viral diseases without causing major toxic side effects. AZT has been used with some success in prolonging the lives of patients with AIDS.
Certain natural products of cells, called interferons, may have potential antiviral and anticancer properties. Interferons are proteins normally synthesized by the cells of vertebrates, including humans, either intrinsically and without stimulation or in response to certain viral infections, chemicals, or immune reactions. In general, the multiplication of viruses is inhibited by interferons, some to a much greater extent than others. Interferons are generally species-specific; i.e., they are effective in inhibiting viral infection only in cells of the same species that naturally synthesize the interferon.
There are three classes of interferons: α-interferons, produced by blood leukocytes; β-interferons, produced by tissue cells and fibroblasts; and γ-interferons (also called immune interferons or interleukins), produced by immune reactions in blood lymphocytes. Interferons are now known to be a subset of a large group of natural cellular substances called cytokines, which signal cells to perform specific functions. Until recently, interferons were difficult to produce commercially because cells and tissues synthesize only small amounts of them. Through recombinant DNA technology, however, large amounts of interferon can be produced.
There has been some success in using interferons to treat viral diseases, such as colds caused by rhinoviruses, infections caused by herpesviruses, and benigntumours and warts caused by papillomaviruses. Local administration at the sites of viral infection affords the best results, although injections of large amounts of interferons can be harmful, probably because they tend to inhibit protein synthesis in the host cell.
Owing to their simplicity, viruses were at first considered to be the primordial life-forms. This concept is almost certainly incorrect, because viruses are completely devoid of the machinery for life processes; therefore, they could not have survived in the absence of cells. Viruses probably evolved from cells rather than cells from viruses. It seems likely that all viruses trace their origins to cellular genes and can be considered as pieces of rogue nucleic acids. Although it is easier to imagine the cellular origin of DNA viruses than RNA viruses, the RNA viruses conceivably could have had their origins from cellular RNA transcripts made from cellular DNA. In fact, the discovery that many cells contain reverse transcriptase capable of converting RNA to DNA seems to suggest that conversion of RNA to DNA and DNA to RNA is not rare. Indeed, some speculate that RNA is the primordial genetic information from which DNA evolved to produce more-complex life-forms.
Other possible progenitors of viruses are the plasmids (small circular DNA molecules independent of chromosomes), which are more readily transferred from cell to cell than are chromosomes. Theoretically, plasmids could have acquired capsid genes, which coded for proteins to coat the plasmid DNA, converting it into a virus.
In brief, it is likely that viruses originated from the degradation of cellular nucleic acids, which acquired the property of being readily transferable from cell to cell during the process of evolution. The fact that normal proto-oncogenes of a cell have nucleic acid sequences that are almost identical to the oncogenes of retroviruses lends credence to the theory that viruses originated from cellular genes.
Viruses that infect animals can jump from one species to another, causing a new, usually severe disease in the new host. For example, in 2003 a virus in the Coronaviridae family jumped from an animal reservoir, believed to be horseshoe bats, to humans, causing a highly pathogenic disease in humans called severe acute respiratory syndrome (SARS). The ability of the SARS coronavirus to jump from horseshoe bats to humans undoubtedly required genetic changes in the virus. The changes are suspected to have occurred in the palmcivet, since the SARS virus present in horseshoe bats is unable to infect humans directly.
Such newly emerged viruses often are highly infectious in humans, since they have not been previously encountered by the human immune system and thus humans have no immunity against them. The coronavirus that caused SARS, for example, spread quickly among humans, becoming a significant disease threat. The virus was quickly brought under control, owing to prohibitions on travel and quarantine measures. In late 2019 another type of coronavirus, called SARS-CoV-2, emerged in China and spread rapidly worldwide, giving rise to a pandemic. SARS-CoV-2 caused an illness known as coronavirus disease 2019 (COVID-19), which was very similar to SARS but caused significantly greater mortality, particularly among persons over age 65.
SARS-CoV-2
The coronavirus SARS-CoV-2, the cause of the COVID-19 pandemic.
Influenza A viruses that infect humans can undergo a dramatic antigenic change, called antigenic shift, which generates viruses that cause pandemics. This dramatic change occurs because influenza A viruses have a large animal reservoir, wild aquatic birds. The RNA genome of influenza A viruses is in the form of eight segments. If an intermediate host, probably the pig, is simultaneously infected with a human and an avian influenza A virus, the genome RNA segments can be reassorted, yielding a new virus that has a surface protein that is immunologically distinct from that of influenza A viruses that have been circulating in the human population. Because the human population will have little or no immunological protection against the new virus, a pandemic will result. This is what most likely occurred in the 1957 flu pandemic, the 1968 flu pandemic, and the influenza pandemic (H1N1) of 2009.
Pandemic influenza A viruses can also apparently arise by a different mechanism. It has been postulated that the strain that caused the influenza epidemic of 1918–19 derived all eight RNA segments from an avian virus and that this virus then underwent multiple mutations in the process of adapting to mammalian cells. The bird flu viruses, which have spread from Asia to Europe and Africa since the 1990s, appear to be taking this route to pandemic capability. These viruses, which have been directly transmitted from chickens to humans, contain only avian genes and are highly pathogenic in humans, causing a mortality rate higher than 50 percent. Bird flu viruses have not yet acquired the ability to transmit efficiently from humans to humans, and it is not known what genetic changes must take place for them to do so.
Viruses are classified on the basis of their nucleic acid content, their size, the shape of their protein capsid, and the presence of a surrounding lipoprotein envelope.
The primary taxonomic division is into two classes based on nucleic acid content: DNA viruses or RNA viruses. The DNA viruses are subdivided into those that contain either double-stranded or single-stranded DNA. The RNA viruses also are divided into those that contain double-stranded or single-stranded RNA. Further subdivision of the RNA viruses is based on whether the RNA genome is segmented. If the viruses contain single-stranded RNA as their genetic information, they are divided into positive-strand viruses if the RNA is of messenger sense (directly translatable into proteins) or negative-strand viruses if the RNA must be transcribed by a polymerase into mRNA.
All viruses falling into one of these nucleic acid classifications are further subdivided on the basis of whether the nucleocapsid (protein coat and enclosed nucleic acid) assumes a rodlike or a polygonal (usually icosahedral) shape. The icosahedral viruses are further subdivided into families on the basis of the number of capsomeres making up the capsids. Finally, all viruses fall into two classes depending on whether the nucleocapsid is surrounded by a lipoprotein envelope.
Some virologists adhere to a division of viruses into those that infect bacteria, plants, or animals; these classifications have some validity, particularly for the unique bacterial viruses with tails, but there is otherwise so much overlap that taxonomy based on hosts seems unworkable. Classification based on diseases caused by viruses also is not tenable, because closely related viruses frequently do not cause the same disease. Eventually, it is likely that the classification of viruses will be based on their nucleotide sequences and their mode of replication rather than on structural components, as is now the case.
The basic taxonomic group is called a family, designated by the suffix -viridae. The major taxonomic disagreement among virologists is whether to segregate viruses within a family into a specific genus and further subdivide them into species names. In the first decade of the 21st century, there occurred a shift toward the use of binomial nomenclature, dividing viruses into italicized genera and species. This move was prompted in large part by the International Committee on Taxonomy of Viruses (ICTV), a member group of the International Union of Microbiological Societies. The ICTV oversees the ongoing process of devising and maintaining a universal classification scheme for viruses. In the virus classification hierarchy, the ICTV recognizes orders, families, subfamilies, genera, and species. The placement of viruses in these groups is based on information provided by study groups composed of experts on specific types of viruses.
In the ICTV system, each species of virus is generally recognized as representing a group of isolates, or viruses with distinct nucleic acid sequences. Thus, a single species of virus may sometimes contain more than one isolate. Although the isolates of a species possess unique genetic sequences, they all descend from the same replicating lineage and therefore share particular genetic traits. Furthermore, isolates of a species also share in common the ability to thrive within a specific ecological niche. As scientists identify new isolates and species, the classification of viruses is expected to become increasingly complex. The following scheme presents examples of well-characterized DNA and RNA viruses as they are classified on the basis of the ICTV system.
Large viruses of complex structure with dimensions of 400 × 250 nm, the genome of which is linear double-stranded DNA. Virions contain at least 40 proteins and lipids, as well as internal structures called lateral bodies. The 2 subfamilies are called Chordopoxvirinae, which infect vertebrates and are closely related antigenically, and Entomopoxvirinae, which infect arthropods. The Chordopoxvirinae are composed of groups called orthopoxviruses (vaccinia), parapoxviruses, avipoxviruses of birds, and many others that infect sheep, rabbits, and swine.
Nonenveloped virions of icosahedral symmetry, about 80 nm in diameter, and capsids containing 252 capsomeres with 12 vertices to which are attached glycoprotein fibres 10–30 nm in length with knobs at the ends. The genome is linear double-stranded DNA. Classified in 2 subgroups: mastadenoviruses, which infect mammals, and aviadenoviruses, which infect birds. Common acute respiratory and gastrointestinal pathogens of humans, and some types cause malignant transformation of cultured cells and can cause cancer in animals.
Icosahedral virions with capsid about 150–200 nm in diameter and 162 capsomeres surrounded by a floppy envelope containing glycoprotein spikes. Genome composed of linear double-stranded DNA. There are 3 known subfamilies: Alphaherpesvirinae, consisting of human herpes simplex viruses types 1 and 2, bovine mamillitis virus, SA8 virus and monkey B virus, pseudorabies virus, equine herpesvirus, and varicella-zoster virus; Betaherpesvirinae, composed of species of cytomegaloviruses; and Gammaherpesvirinae, composed of genera familiarly called Epstein-Barr virus, baboon herpesvirus, chimpanzee herpesvirus, Marek’s disease virus of chickens, turkey herpesvirus, herpesvirus saimiri, and herpesvirus ateles.
Large enveloped or nonenveloped icosahedral virions measuring 120–350 nm in diameter and containing linear double-stranded DNA. Genera include Iridovirus, which contains invertebrate iridescent virus 6, and Lymphocystivirus, which contains lymphocystis disease virus 1 of fish.
Icosahedral, enveloped virions approximately 175–215 nm in diameter that contain linear double-stranded DNA. This family consists of one genus,Asfivirus, which contains the African swine fever virus.
Small enveloped, spherical virions about 40–48 nm in diameter containing circular double-stranded DNA with a single-stranded DNA region and a DNA-dependent DNA polymerase that repairs the single-stranded DNA gap and is essential for replication. Also characteristic are the use of reverse transcriptase for replication and an abundance of a soluble protein (HBsAg). Genera include Orthohepadnavirus, which consists of hepatitis B viruses that infect mammals, and Avihepadnavirus, which consists of hepatitis B viruses that infect birds.
Icosahedral, nonenveloped virions about 52–55 nm in diameter with 72 capsomeres. Virions contain covalently linked circular DNA. Papillomaviruses do not grow in cell culture, and they usually cause warts and benign papillomas; in some instances papillomas develop into cancers. The family contains multiple genera.
Small icosahedral, nonenveloped virions with 32 capsomeres measuring 18–26 nm in diameter that contain single-stranded DNA. Viruses of this family are divided into two subfamilies: Parvovirinae, which infect vertebrates, and Densovirinae, which infect insects. The vertebrate viruses fall into 2 classes: those that replicate autonomously and those that replicate only in the presence of helper adenoviruses or herpesviruses, designated adenoassociated viruses (AAV).
Icosahedral, nonenveloped virions 40–55 nm in diameter. Virions contain covalently linked circular double-stranded DNA. The family consists of one genus, Polyomavirus. The polyomaviruses produce malignant transformation of infected cells.
Small icosahedral, nonenveloped virions 20–30 nm in diameter, composed of 60 capsomeres and containing nonsegmented single-stranded, positive-sense RNA. Among the multiple recognized genera are Enterovirus (polioviruses),Cardiovirus, Rhinovirus (common cold viruses), and Aphthovirus (foot-and-mouth disease virus).
Icosahedral, nonenveloped virions about 35–39 nm in diameter, composed of 32 capsomeres and 180 molecules of a single capsid protein. The genome consists of a single strand of positive-sense RNA. The prototype virus of this family is the vesicular exanthema of swine virus.
Enveloped virions spherical in shape with icosahedral nucleocapsid about 70 nm in diameter. The genome is single-stranded positive-sense RNA. There are 2 recognized genera: Alphavirus and Rubivirus. Alphavirus consists of viruses transmitted by arthropods (exclusively mosquitoes); prototypes include Sindbis virus and eastern and western equine encephalitis viruses. Rubivirus contains non-arthropod-borne viruses, including the causative agent of German measles.
Viruses of this family are enveloped and spherical in shape, with a genome consisting of nonsegmented single-stranded positive-sense RNA. These viruses are transmitted by either insects or arachnids and cause severe diseases such as yellow fever, dengue, tick-borne encephalitis, and Japanese B encephalitis. Other members of this family include non-arthropod-borne hog cholera virus (pestivirus) and hepatitis C virus of humans.
Enveloped virions 120 nm in diameter with a helical nucleocapsid containing a single strand of positive-sense RNA. Club-shaped glycoprotein spikes in envelope give crownlike (coronal) appearance. Viruses of this family are important agents of respiratory and gastrointestinal disease in humans, poultry, and bovines.
Enveloped virions about 80–120 nm in diameter with a helical nucleocapsid containing 8 segments of single-stranded negative-sense RNA and endogenous RNA polymerase. The lipoprotein envelope contains 2 glycoproteins, designated hemagglutinin (major antigen) and neuraminidase. The best-known viruses in this family are the 3 distinct antigenic types of influenza viruses: A, B, and C.
Enveloped virions varying in size from 150 to 200 nm in diameter with a helical nucleocapsid containing a single strand of negative-sense nonsegmented RNA and an endogenous RNA polymerase. The lipoprotein envelope contains 2 glycoprotein spikes designated hemagglutinin-neuraminidase (HN) and fusion factor (F). The major subfamily is Paramyxovirinae, which contains the human parainfluenza viruses and mumps virus, as well as Newcastle disease virus of poultry. The genus Morbillivirus, within Paramyxovirinae, contains the agents that cause measles in humans, distemper in dogs and cats, and rinderpest in cattle. The second subfamily, Pneumovirinae, causes the serious respiratory syncytial virus disease in human infants.
Enveloped virions, usually bullet-shaped, about 75 nm in diameter and 180 nm in length, containing a helical nucleocapsid with single-stranded negative-sense RNA and an endogenous RNA polymerase. The lipoprotein envelope contains a single glycoprotein, which is the type-specific antigen. Viruses of this family are widely infectious for plants and for animals varying from insects to humans. Genera that infect animals are Vesiculovirus, which includes the virus that causes vesicular stomatitis in cattle, swine, and equines, and Lyssavirus, which includes the causative agent of rabies.
Enveloped virions, variably elongated filaments 650–1,400 nm in length and pleomorphic in shape, containing a helical nucleocapsid with single-stranded negative-sense RNA (about 19 kilobases in length) and an endogenous RNA polymerase. Much like the Rhabdoviridae, the lipoprotein envelope contains a single glycoprotein, which is the type-specific antigen. The family consists of 2 genera: Filovirus, which contains the Marburg viruses, and Ebolavirus, which contains the Ebola viruses. These viruses have been isolated from African monkeys, and both are among the most dangerous pathogens. Some strains cause severe hemorrhagic fevers in humans; the mortality rate from these diseases is as high as 90 percent. Human infections with Marburg virus have been traced to laboratory monkeys, but human outbreaks of fatal Ebola virus infection in Congo (Kinshasa) and Sudan have not been traced to monkeys. Instead, these infections are suspected to have been transmitted from fruit-eating bats.
Enveloped virions 110–130 nm in diameter with a helical nucleocapsid in 2 segments containing negative-sense RNA, an endogenous RNA polymerase, and small amounts of ribosomal RNA. The family contains a single genus, Arenavirus, with species widely distributed in animals and causing serious human diseases. Many of these agents are transmitted by insects.
Enveloped virions about 80–120 nm in diameter with a 3-segment helical nucleocapsid containing single-stranded RNA of negative sense and endogenous RNA polymerase. Many viruses grouped in 5 genera: Orthobunyavirus, Phlebovirus, Nairovirus, Tospovirus, and Hantavirus. Most of these viruses are transmitted by arthropods and cause serious human disease.
Enveloped virions about 80–100 nm in diameter with 2 identical copies of single positive-strand RNA in nondefective virions and a reverse transcriptase, which promotes synthesis of double-stranded DNA from the viral RNA template. A hallmark of the virion RNA templates is long terminal repeat (LTR) nucleotide sequences, which serve for integration of the DNA in chromosomes of the host cell. Retroviridae cause cancers in many species of animals, including humans, and are probably derived from normal cell nucleotide sequences called proto-oncogenes. Certain retroviruses of the lentivirus group cause AIDS in humans, monkeys, felines, and cattle.
Nonenveloped icosahedral virions with outer and inner protein shells 60–80 nm in diameter and containing double-stranded RNA in 10 to 12 segments. Viruses in this family infect many species of plants and animals. Reoviridae genera containing species known to infect animals include Orthoreovirus, Orbivirus (widely distributed in insects and vertebrates, including bluetongue disease virus of sheep), Rotavirus (widespread causative agents of gastroenteritis in mammals, including humans), and Cypovirus (prototype causes cytoplasmic polyhedrosis disease in insects).



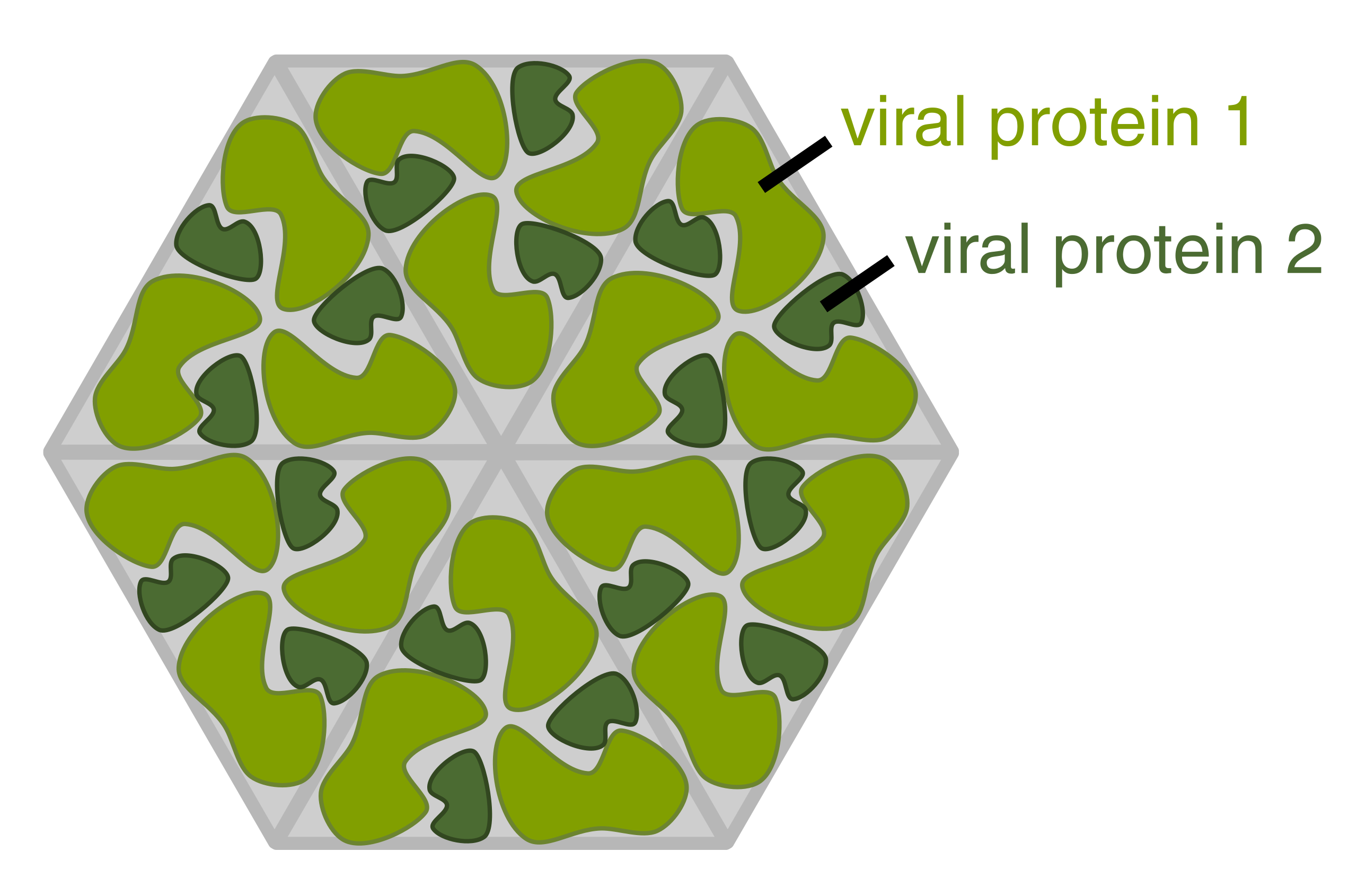
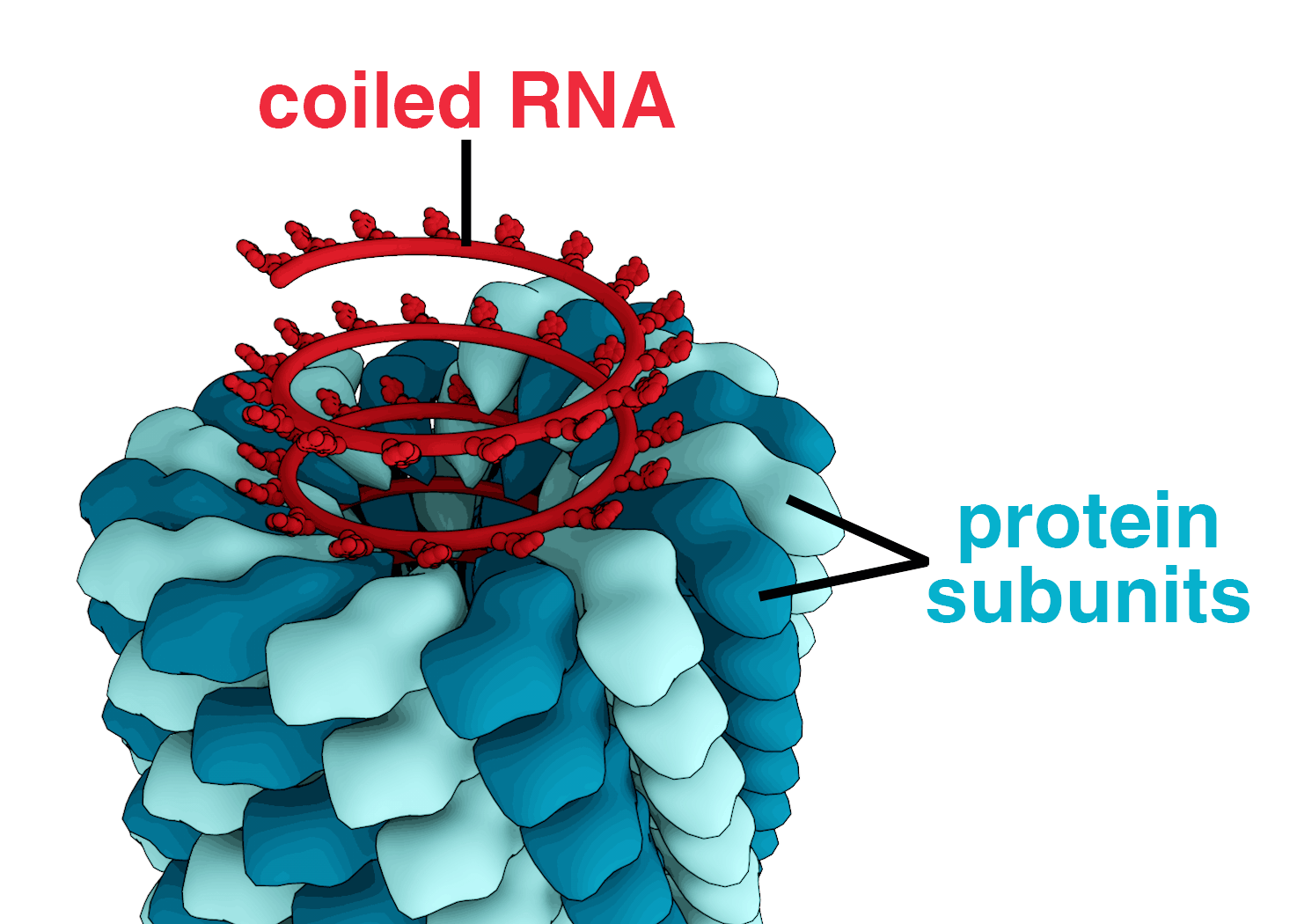

_Virus_PHIL_1878_lores.jpg)