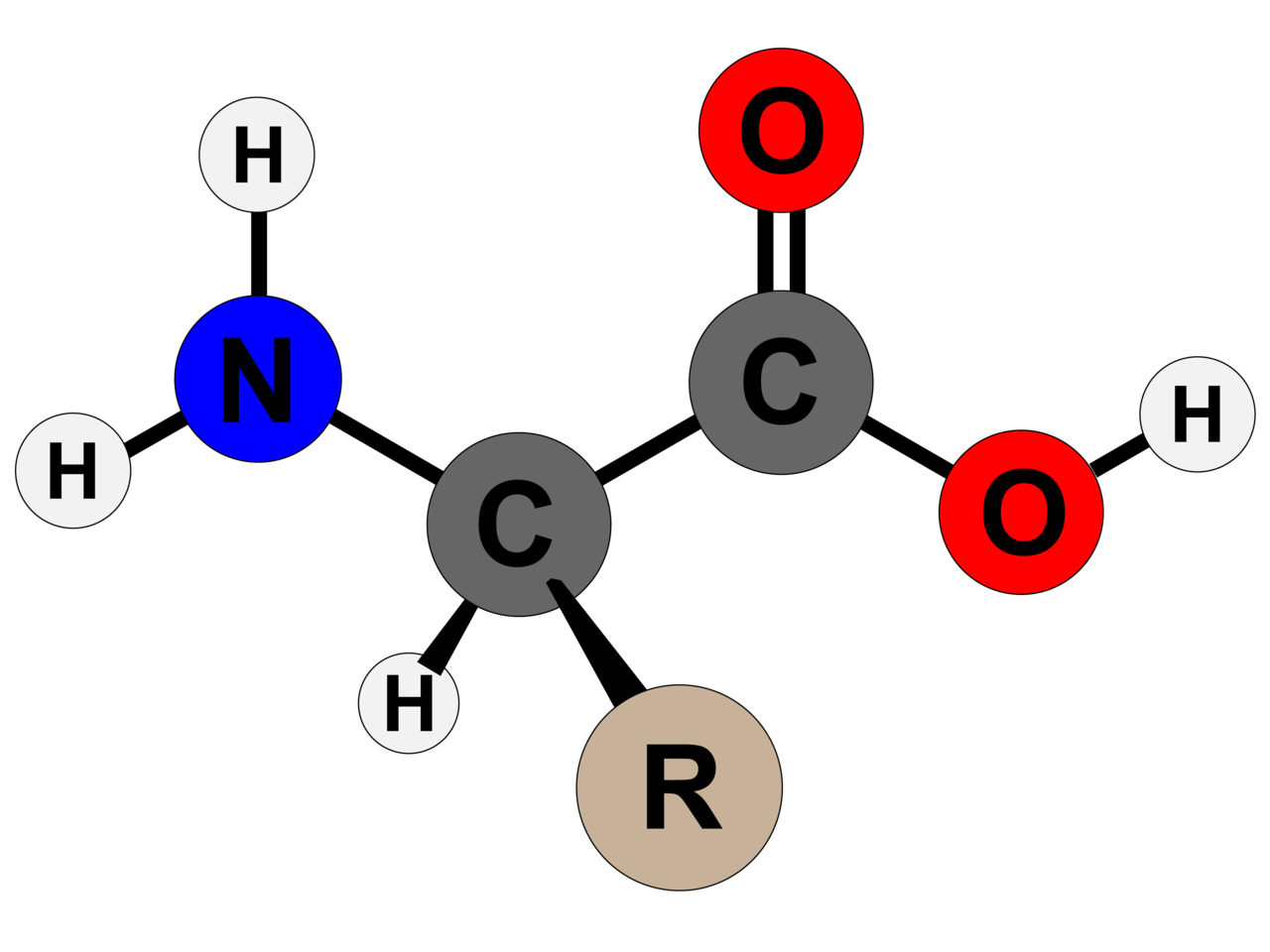
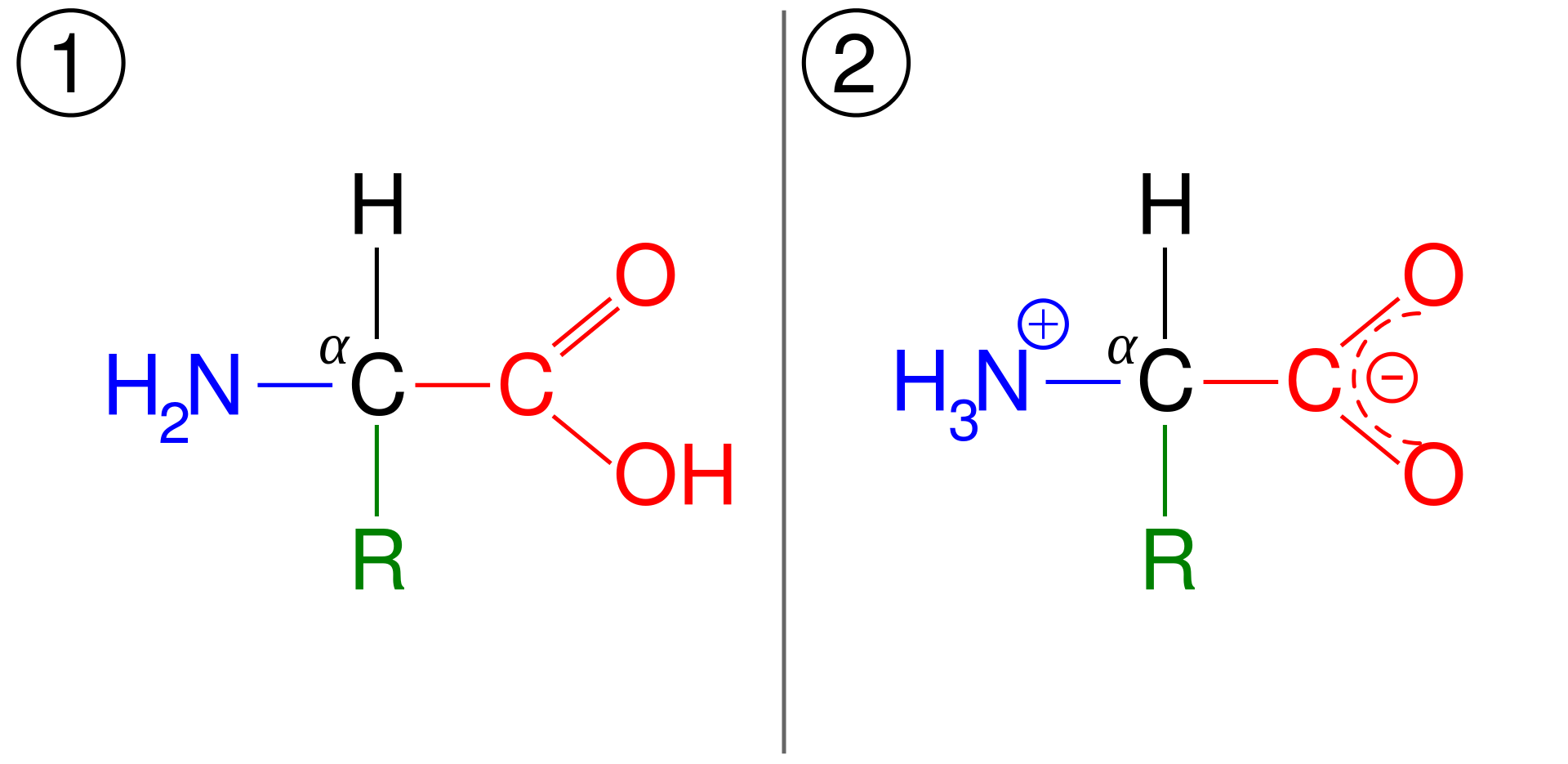
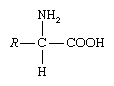Amino asidin genel yapısı
Bu yapı α-amino asitlerden biri (prolin, döngülü bir amino asit) dışında tümüne ortaktır. Karbona bağlı olan R grubu ya da yan zincir her amino asitte ayrıdır.
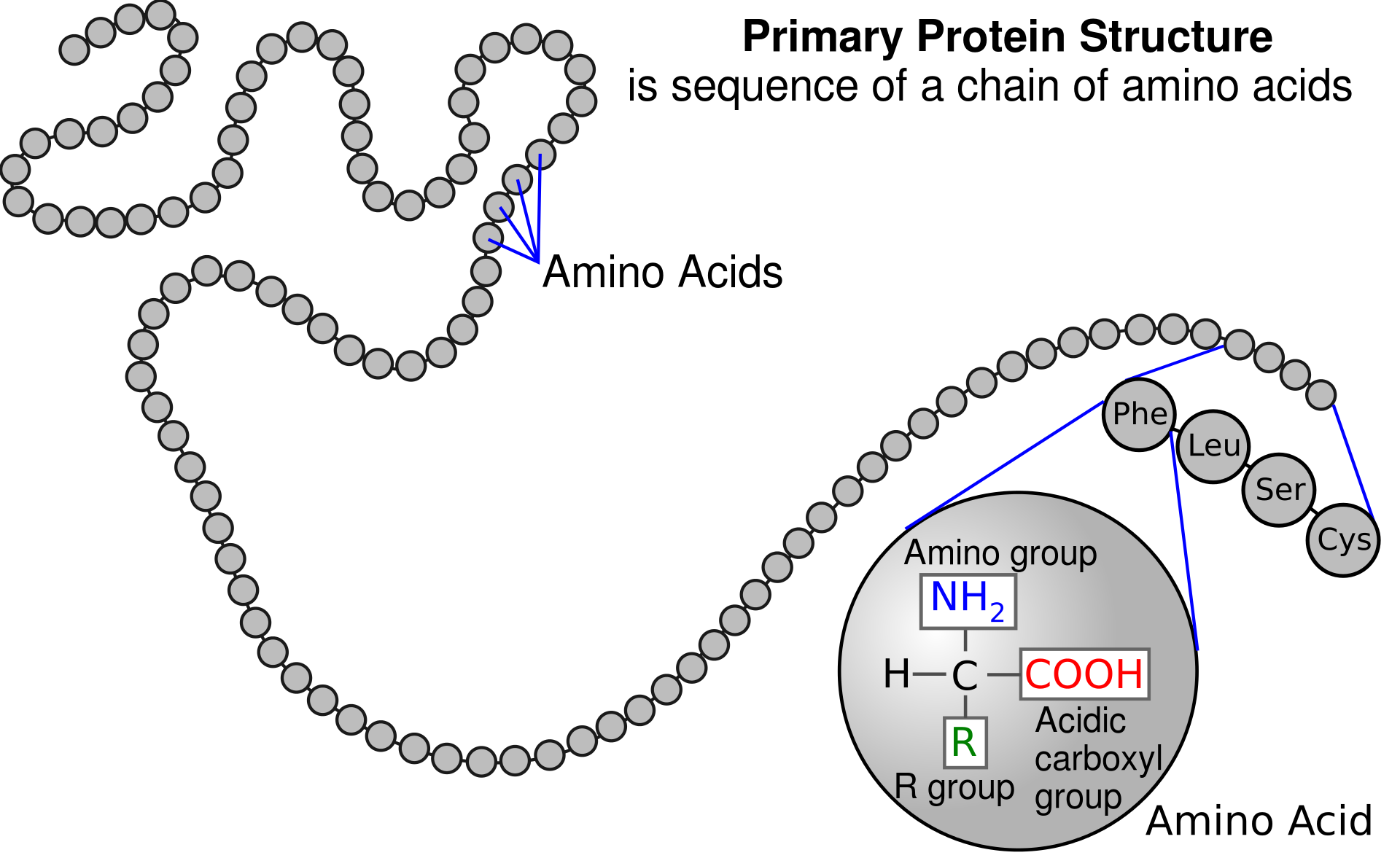
Proteinler kovalent bağ ile bağlı amino asit polimerleridir.
Glysin dışında, tüm ortak ya da ölçün amino asitlerde α-karbon dört ayrı gruba bağlıdır: Bir karboxil grubu (—COOH), bir amino grubu (—NH2), bir R grubu, ve bir H atomu (glysinde R grubu bir başka H atomudur).
“Amino asit” terimi α-amino karboksilik asit için kısaltmadır.
Amino asitlerde kapsanan başlıca elementler C, H, O, ve N’dir.
Doğal olarak oluşan 500 kadar amino asit vardır.
Genetik kodda yalnızca 20 amino asit bulunur.
Bu grupta, 1808’de “asparagine” keşfedilen ilk amino asit ve 1938’de “threonine” keşfedilen son amino asit oldu.
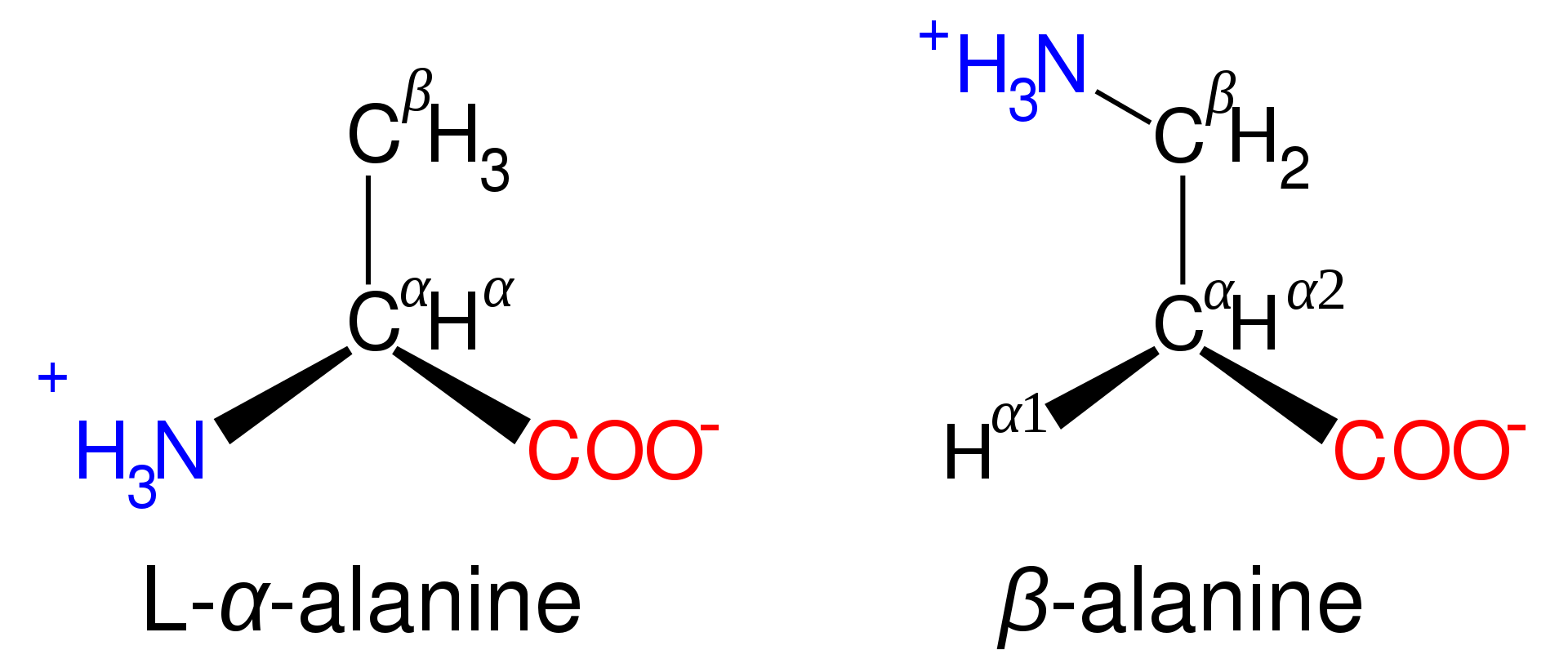
Sınıflandırma: başlıca alfa- (α-), beta- (β-), gamma- (γ-) ya da delta- (δ-) amino asitler olarak yapılır (başka ölçütlere göre de sınıflandırmalar vardır).
Amino asitler bedende proteinler olarak sudan sonra ikinci büyük kütleyi oluşturur.
Amino asitler sinir iletiminde ve biosentezde de işlev görür.
Peptid bağ bir amino asidin amino grubu ile bir başkasının karboksil grubu arasında oluşur.
Kısa polimer zincirlerine peptidler, uzun zincirlere polipeptidler ya da proteinler denir.
Bu zincirler lineer ya da dalsızdır.
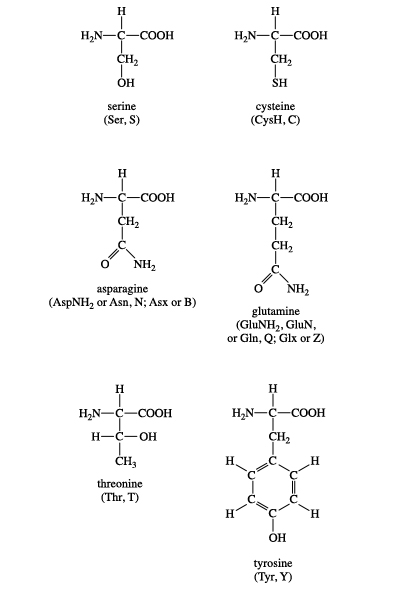
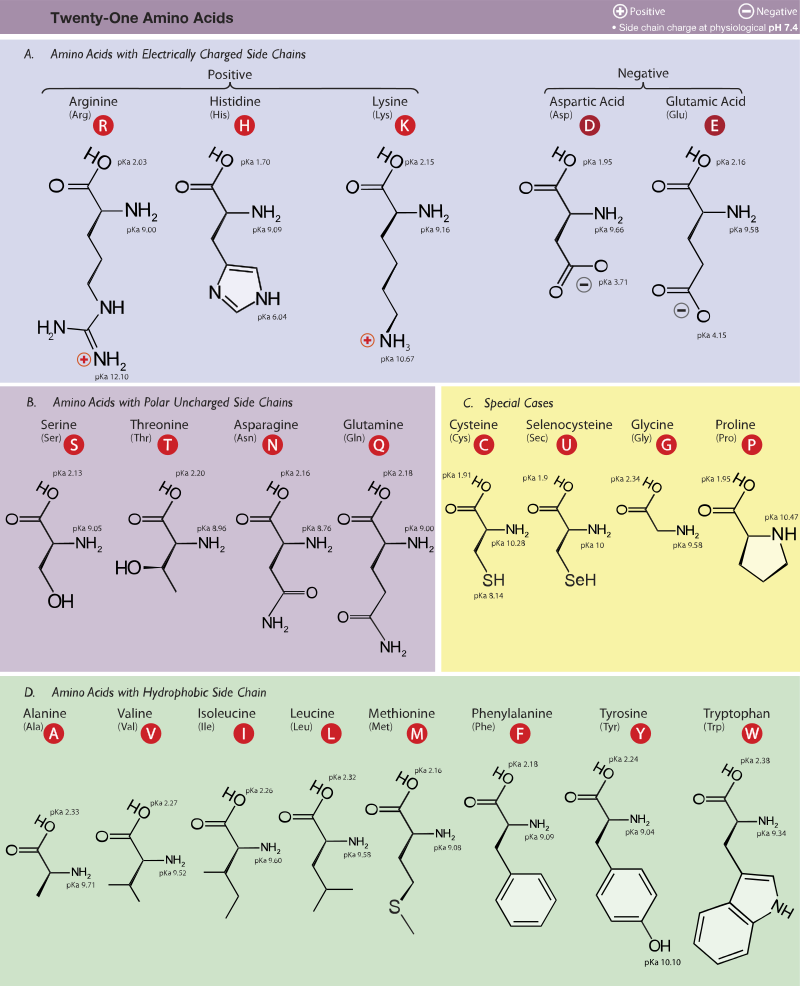
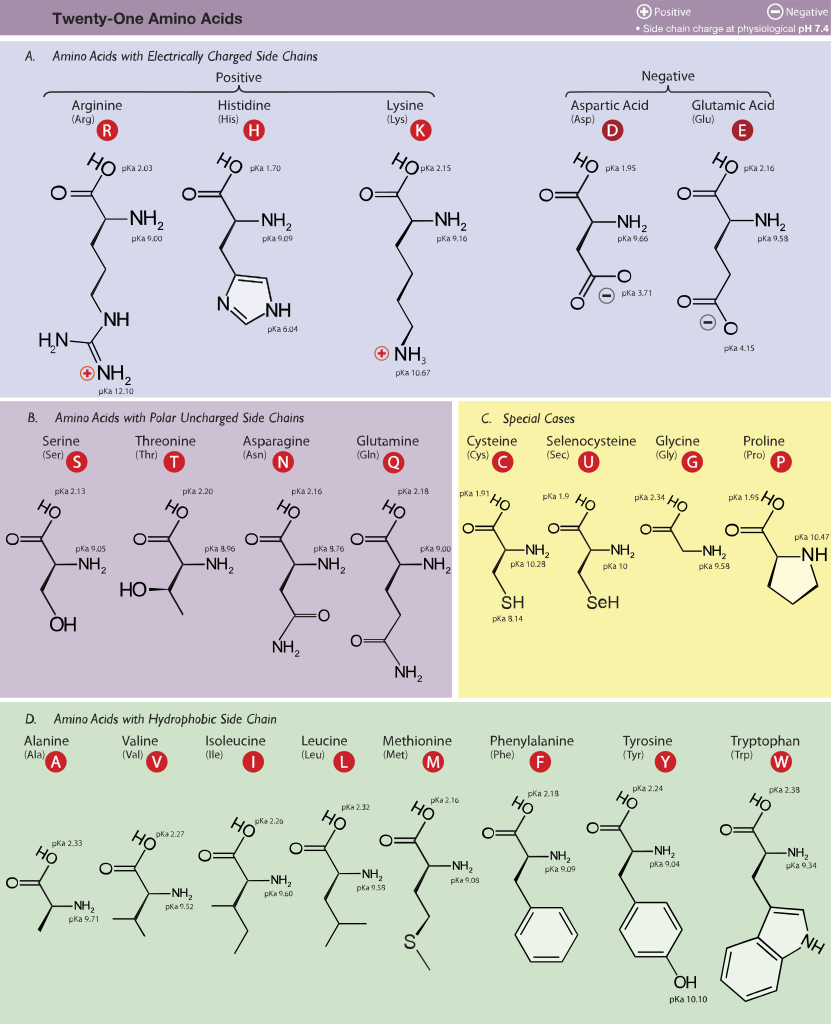
Ökaryotlarda bulunan 21 proteinogenik amino asit. Burada yan zincirlerinin pKa değerlerine ve fizyolojik pHda (7,4) taşıdıkları yüklere göre kümelendirilmişlerdir.
Figür 1.1
Amino asitlerin yapısal özellikleri (tam protonlu biçimlerinde).
Doğada 300’den fazla amino asit türünün olmasına karşın, yalnızca yirmi 20 amino asit DNA tarafından kodlanır ve memeli proteinlerinin bileşenleri olarak bulunur.
Her bir amino asit α-karbon atomuna bağlı bir (asidik işlevli) karboksil grup (—COOH), bir (bazik işlevli) amino grup (—NH2), ve her bir amino aside özgül bir örgensel yan zincirden (R zinciri) oluşur (prolinde R grup Hdir).
Fizyolojik pHda (= 7,4) karboksil grubu ayrılarak negatif yüklü karboksilat ionunu (—COOO-) oluşturur, ve amino grubu protonlanır (—NH3+).
Proteinlerde, bu karboksil ve peptid gruplarının hemen hemen tümü peptid bağ ile bağlıdır ve kimyasal tepkimeye girmez (hidrojen bağ oluşumu dışında).
Böylece, bir amino asidin bir proteinde oynadığı rolü en sonunda yan zincirlerin yapısı belirler.
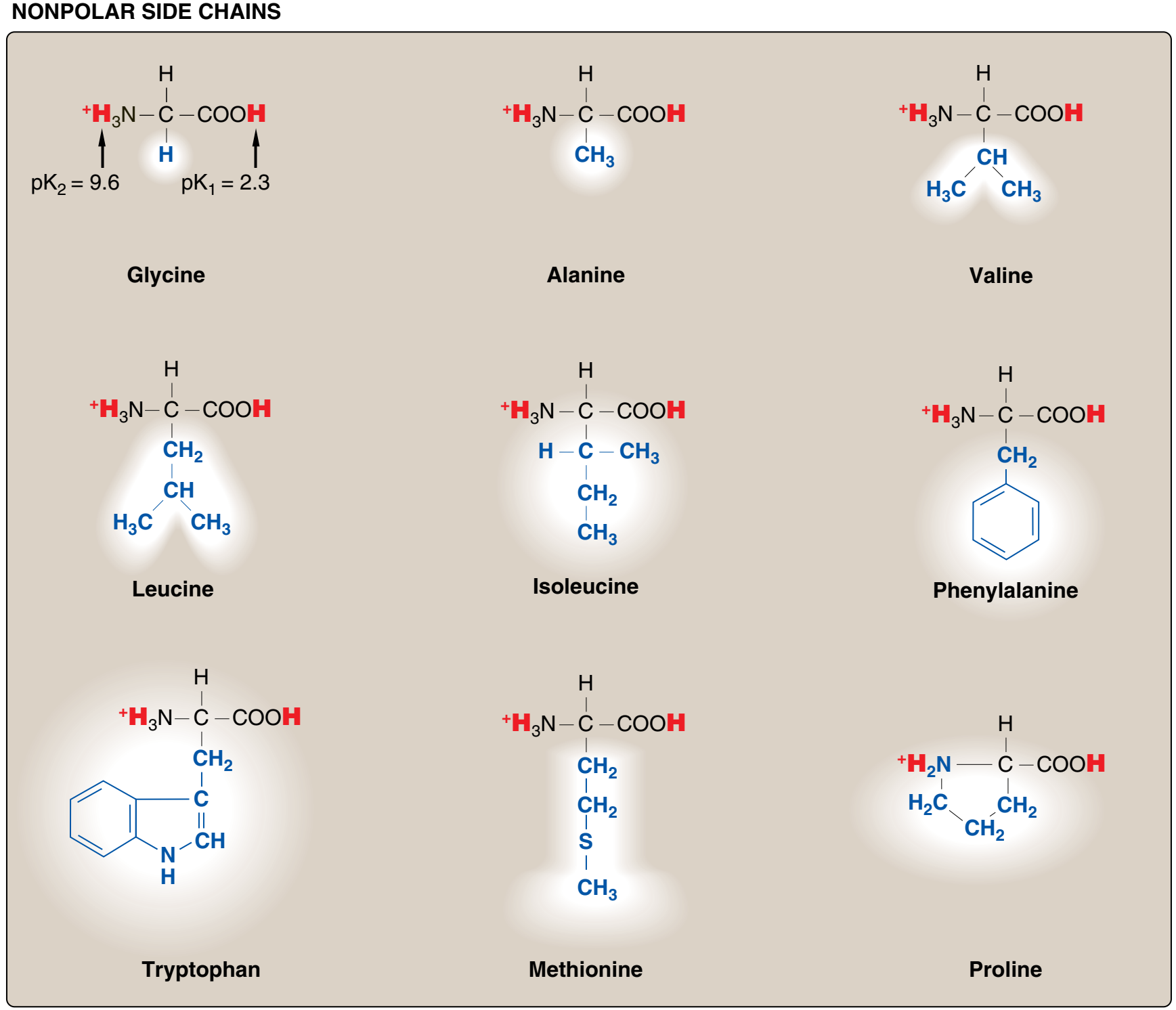
Bu nedenle amino asitleri yan zincirlerinin özelliklerine göre, yani non-polar nonpolar (elektronların dengeli bir dağılımı)
ya da polar (elektronların dengesiz bir dağılımı ile asidik ya da bazik) olmalarına göre sınıflandırmak uygundur (Figürler 1.2 ve 1.3).
Figür 1.2 Proteinlerde ortak olarak bulunan yirmi amino asidin yan zincirlerinin asidik pHda yük ve kutupsallığına göre sınıflandırması (Figür 1.3'te sürüyor).
Her bir amino asit tam protonlanmış biçiminde gösterilmiştir ve ayrılabilir hidrojen ionları kırmızı ile temsil edilir. Non-polar amino asitlerin α-karboksil ve α-amino grupları için pK değerleri glysin için gösterilen değerlere benzerdir.
Figure 1.2
Classification of the 20 amino acids commonly found in proteins, according to the charge and polarity of their
side chains at acidic pH is shown here and continues in Figure 1.3. Each amino acid is shown in its fully protonated
form, with dissociable hydrogen ions represented in red print. The pK values for the α-carboxyl and α-amino
groups of the nonpolar amino acids are similar to those shown for glycine. (Continued in Figure 1.3.)
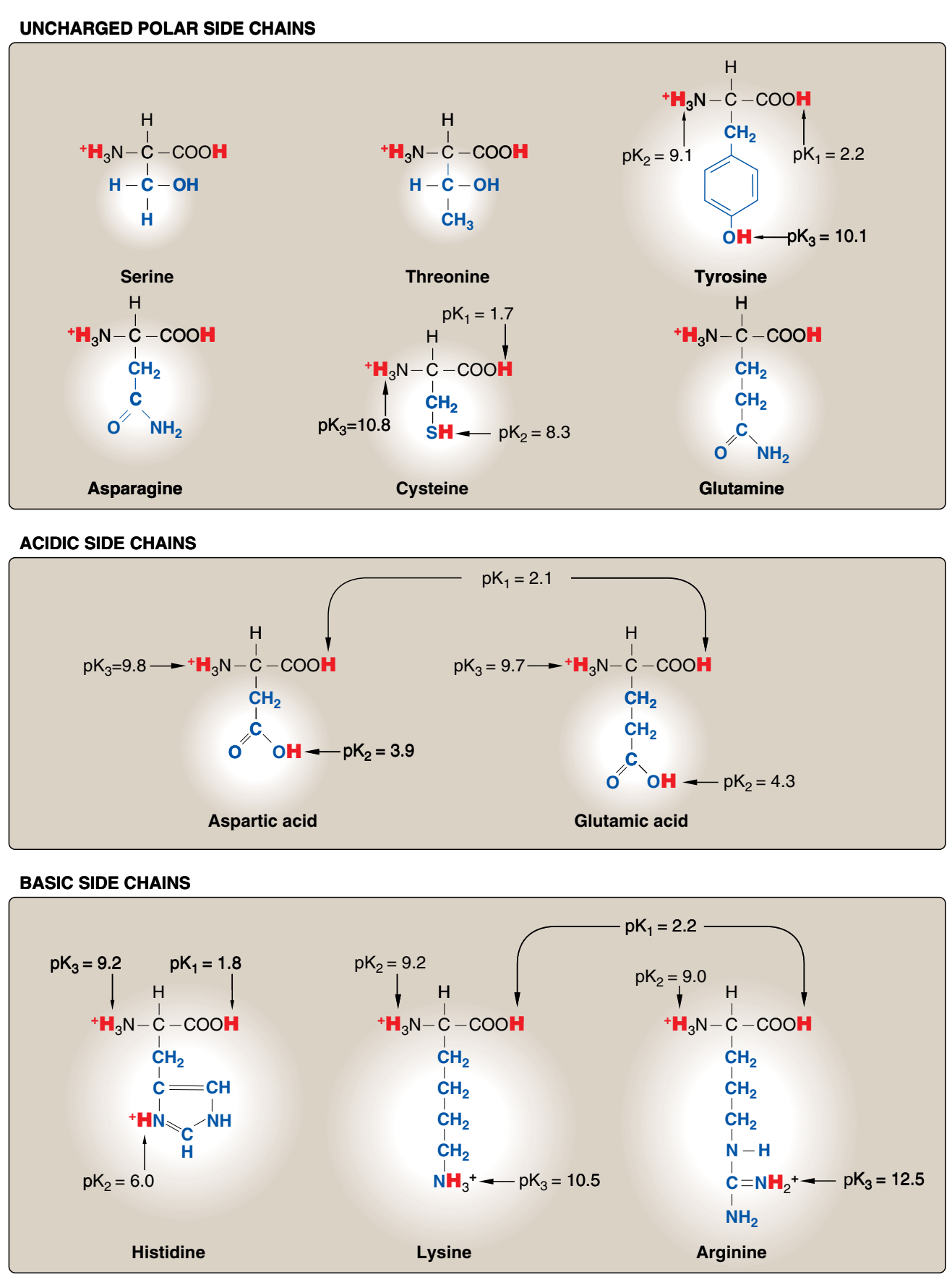
Figür 1.3 Proteinlerde ortak olarak bulunan 20 amino asidin asidik pHda yan zincirlerinin yük ve kutupsallığına göre sınıflandırması (Figür 1.2'den sürüyor).
Figure 1.3
Classification of the 20 amino acids commonly found in proteins, according to the charge and polarity
of their side chains at acidic pH (continued from Figure 1.2)
A. Non-polar yan zincirli amino asitler Bu amino asitlerden her birinin proton kazanmayan ya da yitirmeyen ya da hidrojen ya da ionik bağlara katılmayan bir non-polar yan zinciri vardır (Figür 1.2). Bu amino asitlerin yan zincirleri "yağlı" ya da lipid-benzeri olarak düşünülebilir ve bu özellik hidrofobik tepkimelere götürür.
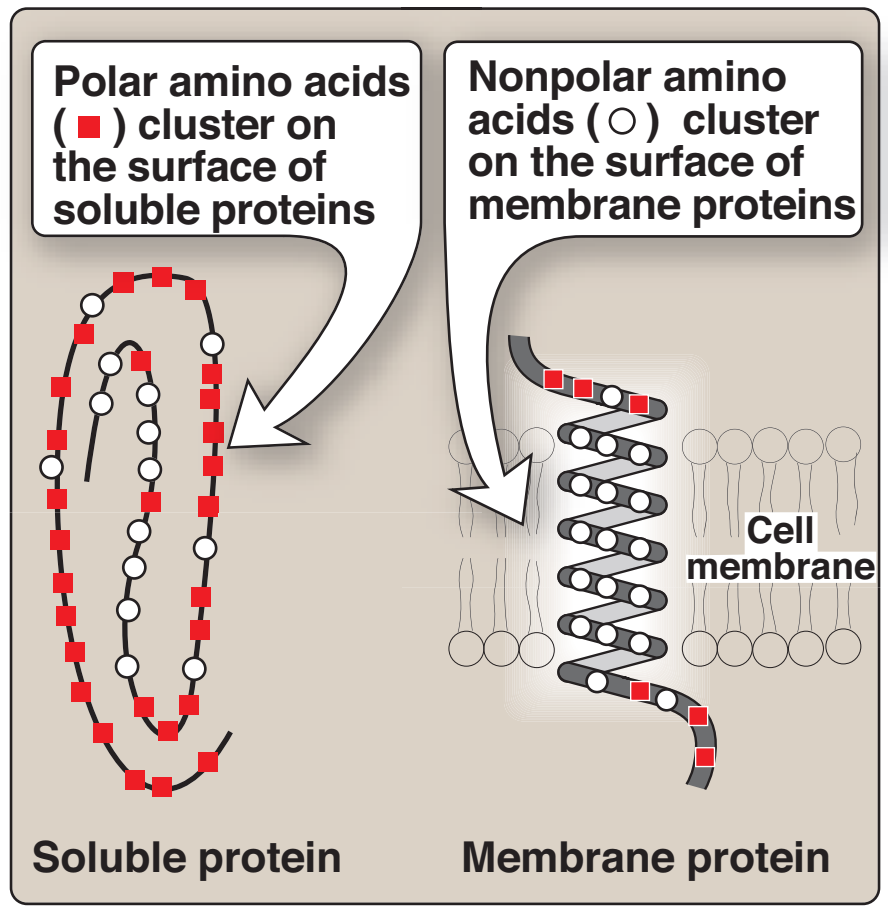
1. Non-polar amino asitlerin konumu: Sulu çözeltilerde — bir polar çevre — bulunan proteinlerde non-polar amino asitlerin yan zincirleri proteinin iç bölgesinde birarada toplanma eğilimindedir (Figür 1.4). Hidrofobik etki olarak bilinen bu fenomen sulu bir çevrede biraraya toplanan yağ damlacıkları gibi davranan non-polar R-gruplarının hidrofobik özelliğine bağlıdır. Non-polar R-gruplar böylece katlanmış proteinin iç bölgesini doldurur ve ona üç-boyutlu şeklini vermede yardımcı olur. Ama örneğin bir membran gibi hidrofobik bir çevrede yerleşmiş olan pnoteinler için non-polar R-gruplar proteinin dış yüzeyinde toplandığı ve lipid çevre ile etkileşim içinde olduğu bulunur (bkz. Figür 1.4). Bu hidrofobik etkileşimler protein yapıyı kararlı kılmada önemlidir.
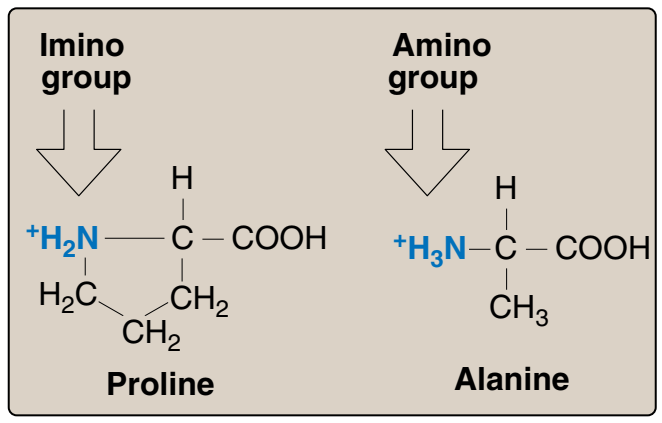
2. Prolin: Prolin yan zincirinin ve α-amino grubunun beş üyeli katı bir halka yapı oluşturmada başka amino asitlerden ayrılır (Figür 1.5). O zaman prolin birincil değil ama ikincil bir amino grubu kapsar. Prolinden sık sık imino asit olarak söz edilir. Prolinin benzersiz geometrisi kolajenin fibril yapısının oluşumuna katkıda bulunur, ve sık sık globüler proteinlerde bulunan α-sarmalları kesintiye uğratır.
A. Amino acids with nonpolar side chains
Each of these amino acids has a nonpolar side chain that does not
gain or lose protons or participate in hydrogen or ionic bonds
(Figure 1.2). The side chains of these amino acids can be thought of
as “oily” or lipid-like, a property that promotes hydrophobic interactions (see Figure 2.10, p. 19).
1. Location of nonpolar amino acids in proteins: In proteins found in
aqueous solutions––a polar environment––the side chains of the
nonpolar amino acids tend to cluster together in the interior of the
protein (Figure 1.4). This phenomenon, known as the hydrophobic effect, is the result of the hydrophobicity of the nonpolar R-groups,
which act much like droplets of oil that coalesce in an aqueous
environment. The nonpolar R-groups thus fill up the interior of the
folded protein and help give it its three-dimensional shape.
However, for proteins that are located in a hydrophobic environment, such as a membrane, the nonpolar R-groups are found on
the outside surface of the protein, interacting with the lipid environment (see Figure 1.4). The importance of these hydrophobic
interactions in stabilizing protein structure is discussed on p. 19.
2. Proline: Proline differs from other amino acids in that proline’s
side chain and α-amino N form a rigid, five-membered ring structure (Figure 1.5). Proline, then, has a secondary (rather than a primary) amino group. It is frequently referred to as an imino acid.
The unique geometry of proline contributes to the formation of the
fibrous structure of collagen (see p. 45), and often interrupts the
α-helices found in globular proteins (see p. 26)
Figure 1.4
Location of nonpolar amino acids
in soluble and membrane proteins.
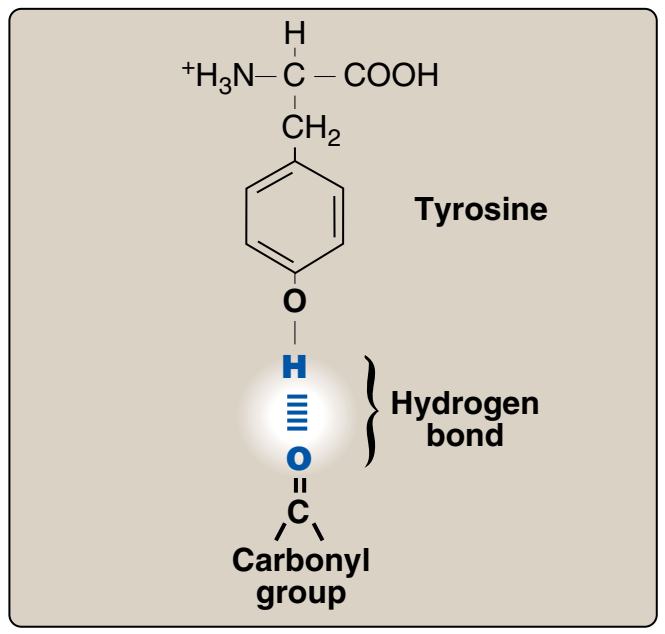
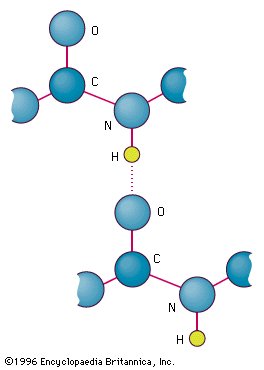
Figür 1.6
Tyrosinin fenolik hidroksil grubu ve bir karboksil grup kapsaan bir başka molekül arasındaki hidrojen bağı.
Figure 1.6
Hydrogen bond between the
phenolic hydroxyl group of tyrosine
and another molecule containing a
carbonyl group.
Figür 1. 5
Prolinde bulunan ikincil amino grubunun örneğin alanin gibi başka amino asitlerde bulunan
birincil amino
grup ile karşılaştırması.
Figure 1.5 Comparison of the secondary
amino group found in proline with
the primary amino group found
in other amino acids, such as
alanine.
B. Yüksüz polar yan zincirli amino asitler Bu amino asitlerin yüksüz pHda sıfır net yükleri vardır, ama cystein ve tyrosinin yan zincirleri alkali bir pHda bir proton yitirebilir (bkz. Figür 1.3). Serin, threonin, ve tyrosin her biri hidrojen bağ oluşumuna katılabilen bir polar hidroksil grup kapsar (Figür 1.6). Asparajin ve glutaminin her birinin yan zincirleri bir karbonil grubu ve bir amid grubu kapsar ve bunların her ikisi de hidrojen bağlarına katılabilir.
1. Disülfid bağ: Systeinin yan zinciri bir sülfhidril grubu (—SH) kapsar ve bu birçok enzimin etkin sitesi için önemli bir bileşendir. Proteinlerde, iki systeinin —SH grubu bir dimer, sistin, oluşturmak üzere oksitlenebilir ve bu bir disülfid bağı (—S—S—) denilen bir kovalent çapraz-bağ kapsar.
2. Başka bileşenler için bağlanma siteleri olarak yan zincirler: Serin, threonin ve seyrek olarak tyrosin polar hidroksil grupları bir fosfat grubu gibi yapılar için bir bağlanma sitesi olarak hizmet edebilir. Ek olarak [Not: Serinin yan zinciri birçok enzimin etkin sitesinin önemli bir bileşenidir.] Ek olarak, asparajinin amid grubu ve ayrıca serin ya da threoninin hidroksil grubu glykoproteinlerde oligosakkaridin bağlanma sitesi olarak hizmet edebilir.
B. Amino acids with uncharged polar side chains
These amino acids have zero net charge at neutral pH, although the
side chains of cysteine and tyrosine can lose a proton at an alkaline
pH (see Figure 1.3). Serine, threonine, and tyrosine each contain a
polar hydroxyl group that can participate in hydrogen bond formation
(Figure 1.6). The side chains of asparagine and glutamine each
contain a carbonyl group and an amide group, both of which can
also participate in hydrogen bonds.
1. Disulfide bond: The side chain of cysteine contains a sulfhydryl
group (—SH), which is an important component of the active site
of many enzymes. In proteins, the –SH groups of two cysteines
can become oxidized to form a dimer, cystine, which contains a
covalent cross-link called a disulfide bond (—S—S—).
2. Side chains as sites of attachment for other compounds: The
polar hydroxyl group of serine, threonine, and, rarely, tyrosine, can
serve as a site of attachment for structures such as a phosphate
group. In addition, the amide group of asparagine, as well as the
hydroxyl group of serine or threonine, can serve as a site of attachment for oligosaccharide chains in glycoproteins.
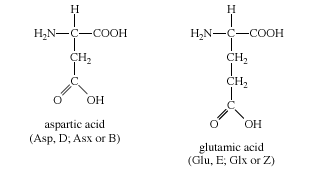
C. Asidik yan zincirleri olan amino asitler Aspartik asit ve glutamik asit amino asitleri proton vericilerdir. Fizyolojik pHda bu amino asitlerin yan zincirleri tam olarak ionlaşır ve negati yüklü bir karboksiklat grubu (—COO-) kapsar. Bu nedenle bu amino asitlere fizyolojik pHda negatif yüklü olduklarını vurgulamak için aspartat ya da glutamat denir (Figür 1.3).
C. Amino acids with acidic side chains
The amino acids aspartic and glutamic acid are proton donors. At
physiologic pH, the side chains of these amino acids are fully ionized,
containing a negatively charged carboxylate group (—COO-) . They are,
therefore, called aspartate or glutamate to emphasize that these amino
acids are negatively charged at physiologic pH (see Figure 1.3).
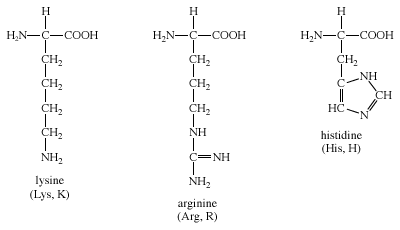
D. Bazik yan zincirli amino asitler Bazik amino asitlerin yan zincirleri proton kabul eder (bkz. Figür 1.3). Fizyolojik pHda lysin ve argininin yan zincirleri tam olarak ionlaşır ve pozitif olarak yüklenir. Karşıt olarak, histidin zayıf olarak baziktir, ve serbest amino asit fizyolojik pHda büyük ölçüde yüksüzdür. Ama histidin bir proteine katıldığı zaman, yan zinciri ya pozitif yüklü ya da yüksüz olabilir ve bu durum proteinin polipeptid zincirlerinin sağladığı ionik çevreye bağımlıdır. Bu histidinin önemli bir özelliğidir ve örneğin hemoglobin gibi proteinlerin işlev görmesinde oynadığı role katkıda bulunur.
D. Amino acids with basic side chains
The side chains of the basic amino acids accept protons (see Figure
1.3). At physiologic pH the side chains of lysine and arginine are fully
ionized and positively charged. In contrast, histidine is weakly basic,
and the free amino acid is largely uncharged at physiologic pH.
However, when histidine is incorporated into a protein, its side chain
can be either positively charged or neutral, depending on the ionic
environment provided by the polypeptide chains of the protein. This
is an important property of histidine that contributes to the role it
plays in the functioning of proteins such as hemoglobin.
📘 Molecular Biology of the Cell (Sixth Edition)
📹📹📹 Amino Acid (VİDEO)
📹 Introduction to Amino Acids / The Organic Chemistry Tutor (VİDEO)
📹 Introduction to Amino Acids / The Organic Chemistry Tutor (LINK)
This biochemistry video tutorial provides a basic introduction into amino acids. It discusses what to look for when describing the side chain groups as polar, nonpolar, acidic, basic, or aromatic.
📹 The 20 Amino Acids / The Organic Chemistry Tutor (VİDEO)
📹 The 20 Amino Acids / The Organic Chemistry Tutor (LINK)
In this video, you'll learn about the general structure of amino acids, which include an amine group, a carboxylic acid group, and a unique side chain (R-group) attached to the alpha carbon. In addition, we'll explore where amino acids fit in terms of biological metabolism.
📹 Amino Acids Intro to Structure Charge Classification and Reactions (VİDEO)
📹 Amino Acids Intro to Structure Charge Classification and Reactions (LINK)
Introduction to Amino Acids on the MCAT including backbone and side chain classification.
Proteinler genlerdeki DNA kodlarının amino asit zincirlerine (üstte) çevrilmesi yoluyla yapılır ve bu amino asit zincirleri üç boyutlu proteinlere (altta) katlanır. Hücrede her bir protein hemen kendine uygun şekle, en düşük enerjili yapıya girer. Katlanma sürecinde içerilen tüm değişkenler — amino asitlerin uzunluğu ve dizilişi, oluşturdukları bağların açıları vb. — hesaplandığı zaman, bir proteinin olanaklı katlanma yollarının sayısı astronomiktir. (L)
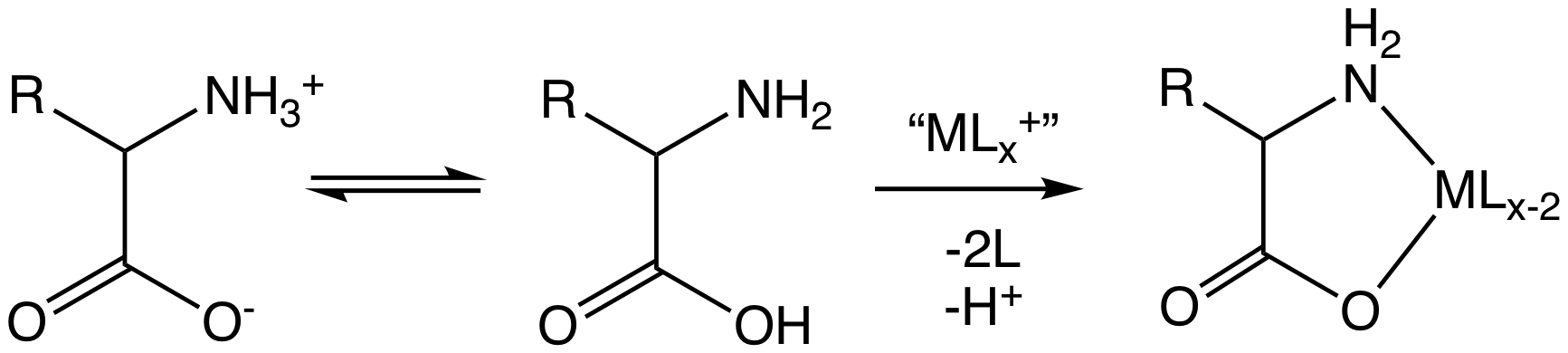
Amino acid, any of a group of organic molecules that consist of a basic amino group (―NH2), an acidic carboxyl group (―COOH), and an organic R group (or side chain) that is unique to each amino acid. The term amino acid is short for α-amino [alpha-amino] carboxylic acid. Each molecule contains a central carbon (C) atom, called the α-carbon, to which both an amino and a carboxyl group are attached. The remaining two bonds of the α-carbon atom are generally satisfied by a hydrogen (H) atom and the R group. The formula of a general amino acid is:
The amino acids differ from each other in the particular chemical structure of the R group.
Proteins are of primary importance to the continuing functioning of life on Earth. Proteins catalyze the vast majority of chemical reactions that occur in the cell. They provide many of the structural elements of a cell, and they help to bind cells together into tissues. Some proteins act as contractile elements to make movement possible. Others are responsible for the transport of vital materials from the outside of the cell (“extracellular”) to its inside (“intracellular”). Proteins, in the form of antibodies, protect animals from disease and, in the form of interferon, mount an intracellular attack against viruses that have eluded destruction by the antibodies and other immune system defenses. Many hormones are proteins. Last but certainly not least, proteins control the activity of genes (“gene expression”).
This plethora of vital tasks is reflected in the incredible spectrum of known proteins that vary markedly in their overall size, shape, and charge. By the end of the 19th century, scientists appreciated that, although there exist many different kinds of proteins in nature, all proteins upon their hydrolysis yield a class of simpler compounds, the building blocks of proteins, called amino acids. The simplest amino acid is called glycine,named for its sweet taste (glyco, “sugar”). It was one of the first amino acids to be identified, having been isolated from the protein gelatin in 1820. In the mid-1950s scientists involved in elucidating the relationship between proteins and genes agreed that 20 amino acids (called standard or common amino acids) were to be considered the essential building blocks of all proteins. The last of these to be discovered, threonine, had been identified in 1935.
All the amino acids but glycine are chiral molecules. That is, they exist in two optically active asymmetric forms (called enantiomers) that are the mirror images of each other. (This property is conceptually similar to the spatial relationship of the left hand to the right hand.) One enantiomer is designated D and the other L. It is important to note that the amino acids found in proteins almost always possess only the L-configuration. This reflects the fact that the enzymes responsible for protein synthesis have evolved to utilize only the L-enantiomers. Reflecting this near universality, the prefix L is usually omitted. Some D-amino acids are found in microorganisms, particularly in the cell walls of bacteria and in several of the antibiotics. However, these are not synthesized in the ribosome.
Another important feature of free amino acids is the existence of both a basic and an acidic group at the α-carbon. Compounds such as amino acids that can act as either an acid or a base are called amphoteric. The basic amino group typically has a pKa between 9 and 10, while the acidic α-carboxyl group has a pKa that is usually close to 2 (a very low value for carboxyls). The pKa of a group is the pH value at which the concentration of the protonated group equals that of the unprotonated group. Thus, at physiological pH (about 7–7.4), the free amino acids exist largely as dipolar ions or “zwitterions” (German for “hybrid ions”; a zwitterion carries an equal number of positively and negatively charged groups). Any free amino acid and likewise any protein will, at some specific pH, exist in the form of a zwitterion. That is, all amino acids and all proteins, when subjected to changes in pH, pass through a state at which there is an equal number of positive and negative charges on the molecule. The pH at which this occurs is known as the isoelectric point (or isoelectric pH) and is denoted as pI. When dissolved in water, all amino acids and all proteins are present predominantly in their isoelectric form. Stated another way, there is a pH (the isoelectric point) at which the molecule has a net zero charge (equal number of positive and negative charges), but there is no pH at which the molecule has an absolute zero charge (complete absence of positive and negative charges). That is, amino acids and proteins are always in the form of ions; they always carry charged groups. This fact is vitally important in considering further the biochemistry of amino acids and proteins.
One of the most useful manners by which to classify the standard (or common) amino acids is based on the polarity (that is, the distribution of electric charge) of the R group (e.g., side chain).
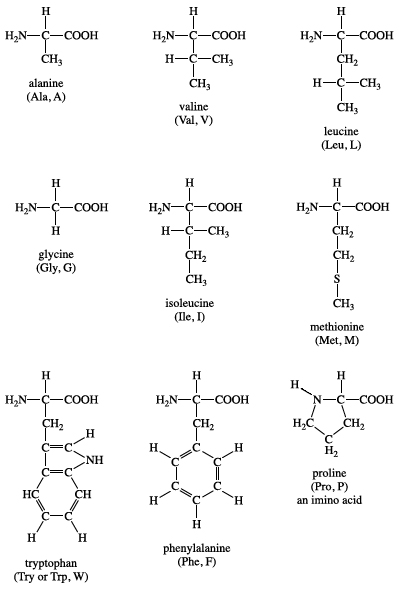
Isoleucine is an isomer of leucine, and it contains two chiral carbon atoms. Proline is unique among the standard amino acids in that it does not have both free α-amino and free α-carboxyl groups. Instead, its side chain forms a cyclic structure as the nitrogen atom of proline is linked to two carbon atoms. (Strictly speaking, this means that proline is not an amino acid but rather an α-imino acid.) Phenylalanine, as the name implies, consists of a phenyl group attached to alanine. Methionine is one of the two amino acids that possess a sulfur atom. Methionine plays a central role in protein biosynthesis (translation) as it is almost always the initiating amino acid. Methionine also provides methyl groups for metabolism. Tryptophan contains an indole ring attached to the alanyl side chain.
Two amino acids, serine and threonine, contain aliphatic hydroxyl groups (that is, an oxygen atom bonded to a hydrogen atom, represented as ―OH). Tyrosine possesses a hydroxyl group in the aromatic ring, making it a phenol derivative. The hydroxyl groups in these three amino acids are subject to an important type of posttranslational modification: phosphorylation (see belowNonstandard amino acids). Like methionine, cysteine contains a sulfur atom. Unlike methionine’s sulfur atom, however, cysteine’s sulfur is very chemically reactive (see belowCysteine oxidation). Asparagine, first isolated from asparagus, and glutamine both contain amideR groups. The carbonyl group can function as a hydrogen bond acceptor, and the amino group (NH2) can function as a hydrogen bond donor.
The two amino acids in this group are aspartic acid and glutamic acid. Each has a carboxylic acid on its side chain that gives it acidic (proton-donating) properties. In an aqueous solution at physiological pH, all three functional groups on these amino acids will ionize, thus giving an overall charge of −1. In the ionic forms, the amino acids are called aspartate and glutamate. The chemical structures of Group III amino acids are
The side chains of aspartate and glutamate can form ionic bonds (“salt bridges”), and they can also function as hydrogen bond acceptors. Many proteins that bind metal ions (“metalloproteins”) for structural or functional purposes possess metal-binding sites containing aspartate or glutamate side chains or both. Free glutamate and glutamine play a central role in amino acid metabolism. Glutamate is the most abundant excitatory neurotransmitter in the central nervous system.
The three amino acids in this group are arginine,histidine, and lysine. Each side chain is basic (i.e., can accept a proton). Lysine and arginine both exist with an overall charge of +1 at physiological pH. The guanidino group in arginine’s side chain is the most basic of all R groups (a fact reflected in its pKa value of 12.5). As mentioned above for aspartate and glutamate, the side chains of arginine and lysine also form ionic bonds. The chemical structures of Group IV amino acids are
The imidazole side chain of histidine allows it to function in both acid and base catalysis near physiological pH values. None of the other standard amino acids possesses this important chemical property. Therefore, histidine is an amino acid that most often makes up the active sites of protein enzymes.
The majority of amino acids in Groups II, III, and IV are hydrophilic (“water loving”). As a result, they are often found clustered on the surface of globular proteins in aqueous solutions.
Amino acids via their various chemical functionalities (carboxyls, amino, and R groups) can undergo numerous chemical reactions. However, two reactions (peptide bond and cysteine oxidation) are of particular importance because of their effect on protein structure.
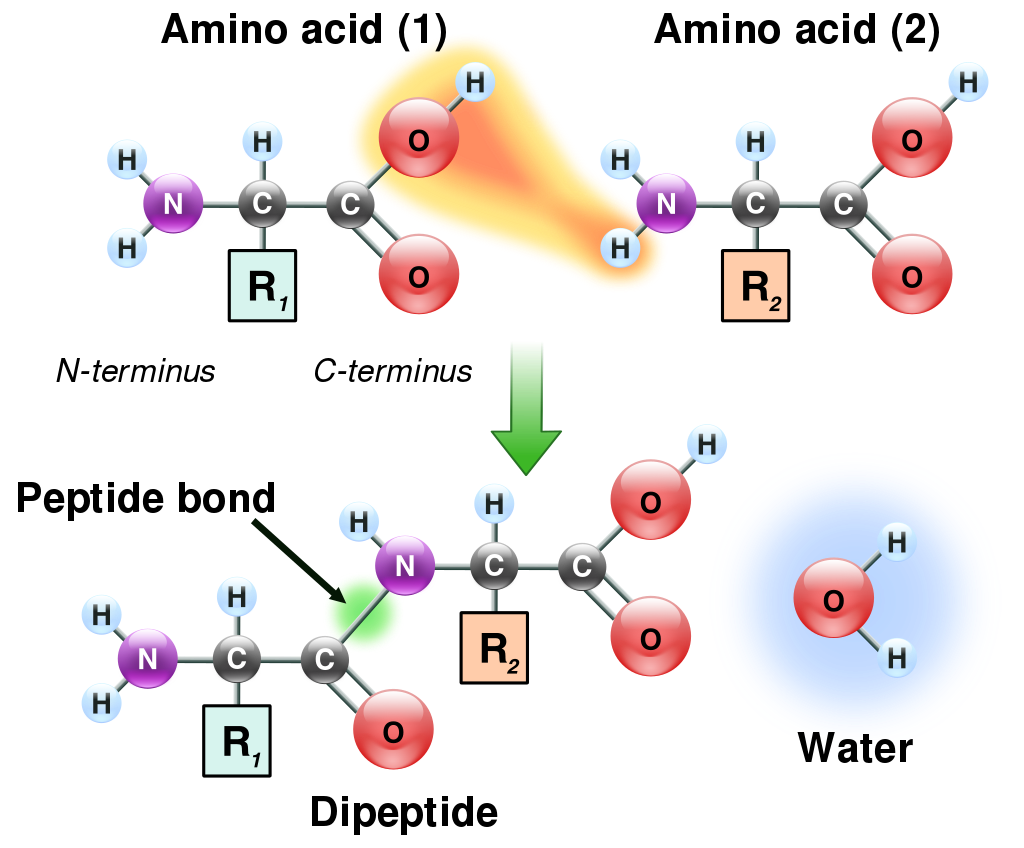
Amino acids can be linked by a condensation reaction in which an ―OH is lost from the carboxyl group of one amino acid along with a hydrogen from the amino group of a second, forming a molecule of water and leaving the two amino acids linked via an amide—called, in this case, a peptide bond. At the turn of the 20th century, German chemist Emil Fischer first proposed this linking together of amino acids. Note that when individual amino acids are combined to form proteins, their carboxyl and amino groups are no longer able to act as acids or bases, since they have reacted to form the peptide bond. Therefore, the acid-base properties of proteins are dependent upon the overall ionization characteristics of the individual R groups of the component amino acids.
The linking of atoms in a peptide bond..
Amino acids joined by a series of peptide bonds are said to constitutea peptide. After they are incorporated into a peptide, the individual amino acids are referred to as amino acid residues. Small polymers of amino acids (fewer than 50) are called oligopeptides, while larger ones (more than 50) are referred to as polypeptides. Hence, a protein molecule is a polypeptide chain composed of many amino acid residues, with each residue joined to the next by a peptide bond. The lengths for different proteins range from a few dozen to thousands of amino acids, and each protein contains different relative proportions of the 20 standard amino acids.
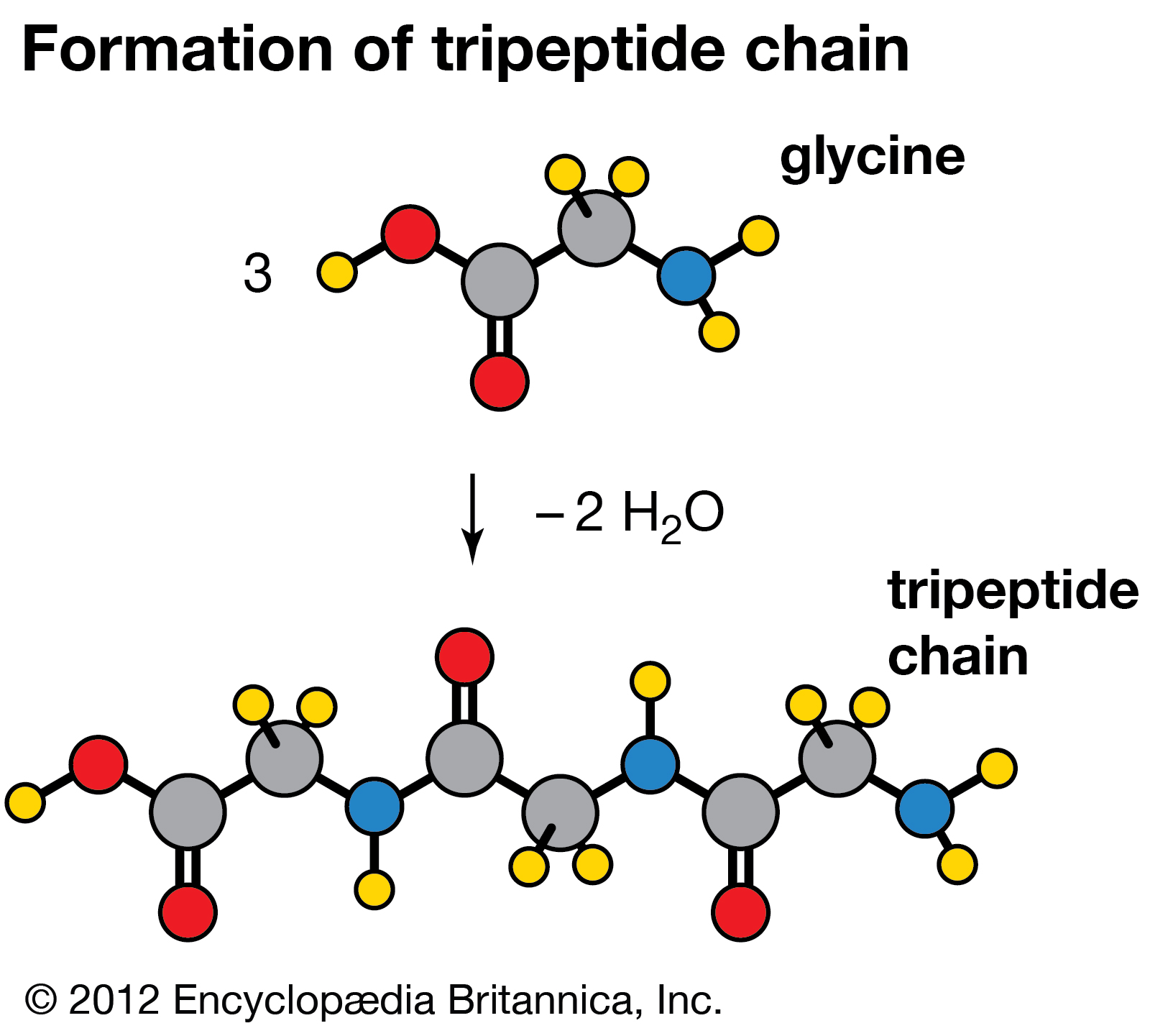
Condensation reaction in which three molecules of the amino acid glycine produce a tripeptide chain, with the elimination of two molecules of water (H2O).
The thiol (sulfur-containing) group of cysteine is highly reactive. The most common reaction of this group is a reversible oxidation that forms a disulfide. Oxidation of two molecules of cysteine forms cystine, a molecule that contains a disulfide bond. When two cysteine residues in a protein form such a bond, it is referred to as a disulfide bridge. Disulfide bridges are a common mechanism used in nature to stabilize many proteins. Such disulfide bridges are often found among extracellular proteins that are secreted from cells. In eukaryotic organisms, formation of disulfide bridges occurs within the organelle called the endoplasmic reticulum.
In extracellular fluids (such as blood), the sulfhydryl groups of cysteine are rapidly oxidized to form cystine. In a genetic disorder known as cystinuria, there is a defect that results in excessive excretion of cystine into the urine. Because cystine is the least soluble of the amino acids, crystallization of the excreted cystine results in formation of calculi—more commonly known as “stones”—in the kidney, ureter, or urinary bladder. The stones may cause intense pain, infection, and blood in the urine. Medical intervention often involves the administration of D-penicillamine. Penicillamine works by forming a complex with cystine; this complex is 50 times more water-soluble than cystine alone.
In summary, it is the sequence of amino acids that determines the shape and biological function of a protein as well as its physical and chemical properties. Thus, the functional diversity of proteins arises because proteins are polymers of 20 different kinds of amino acids. For example, a “simple” protein is the hormoneinsulin, which has 51 amino acids. With 20 different amino acids to chose from at each of these 51 positions, a total of 2051, or about 1066, different proteins could theoretically be made.
polynucleotide chain of deoxyribonucleic acid (DNA)
Portion of polynucleotide chain of deoxyribonucleic acid (DNA). The inset shows the corresponding pentose sugar and pyrimidine base in ribonucleic acid (RNA).
Several standard and nonstandard amino acids often are vital metabolic intermediates. Important examples of this are the amino acids arginine, citrulline, and ornithine, which are all components of the urea cycle. The synthesis of urea is the principal mechanism for the removal of nitrogenous waste.
Nonstandard amino acids refer to those amino acids that have been chemically modified after they have been incorporated into a protein (called a “posttranslational modification”) and those amino acids that occur in living organisms but are not found in proteins. Among these modified amino acids is γ-carboxyglutamic acid, a calcium-binding amino acid residue found in the blood-clotting protein prothrombin (as well as in other proteins that bind calcium as part of their biological function). The most abundant protein by mass in vertebrates is collagen. Significant proportions of the amino acids in collagen are modified forms of proline and lysine: 4-hydroxyproline and 5-hydroxylysine.
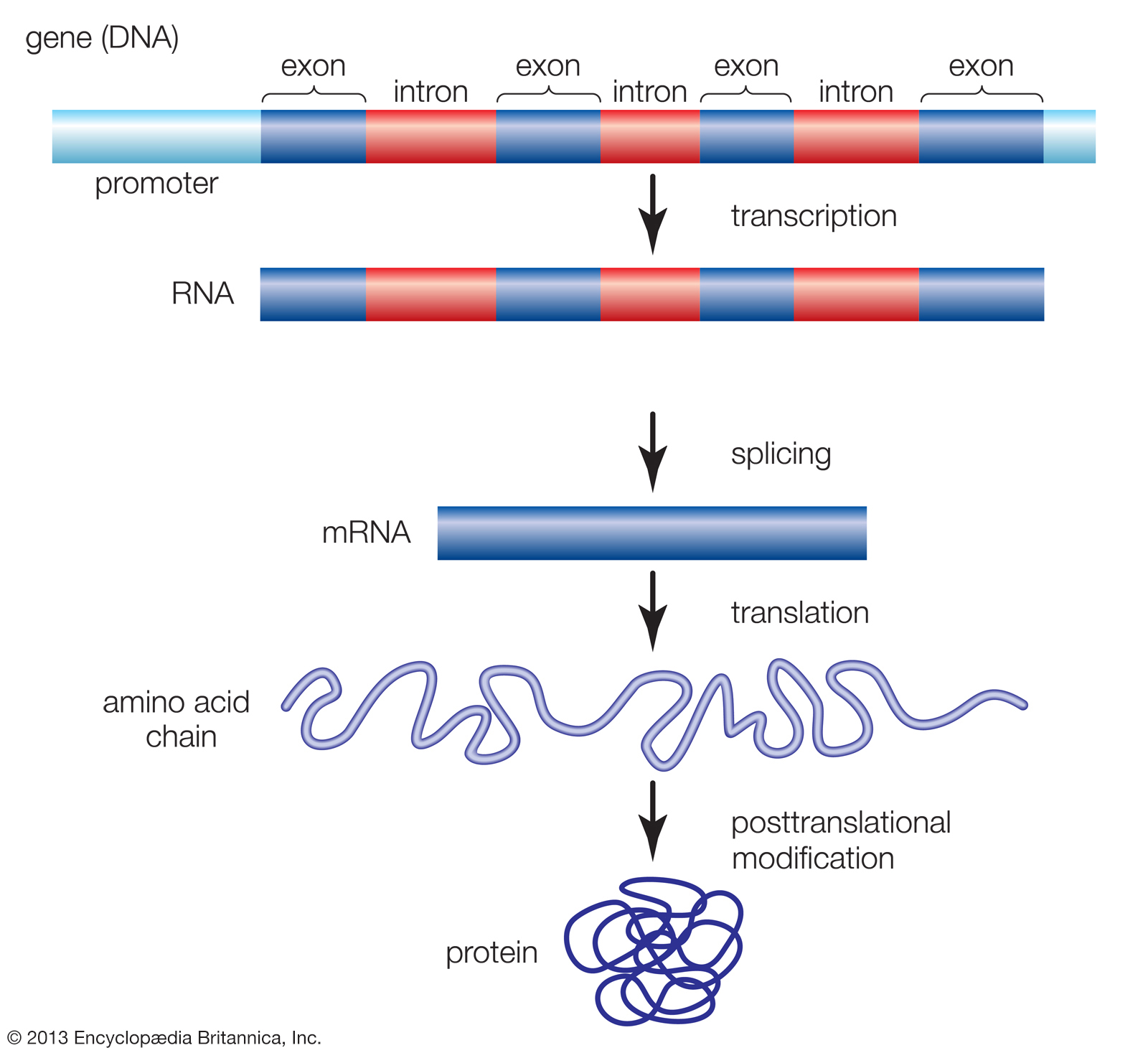
Genes are made up of promoter regions and alternating regions of introns (noncoding sequences) and exons (coding sequences). The production of a functional protein involves the transcription of the gene from DNA into RNA, the removal of introns and splicing together of exons, the translation of the spliced RNA sequences into a chain of amino acids, and the posttranslational modification of the protein molecule.
Arguably, the most important posttranslational modification of amino acids in eukaryotic organisms (including humans) is the reversible addition of a phosphate molecule to the hydroxyl portion of the R groups of serine,threonine, and tyrosine. This event is known as phosphorylation and is used to regulate the activity of proteins in their minute-to-minute functioning in the cell. Serine is the most commonly phosphorylated residue in proteins, threonine is second, and tyrosine is third.
Proteins with carbohydrates (sugars) covalently attached to them are called glycoproteins. Glycoproteins are widely distributed in nature and provide the spectrum of functions already discussed for unmodified proteins. The sugar groups in glycoproteins are attached to amino acids through either oxygen (O-linked sugars) or nitrogen atoms (N-linked sugars) in the amino acid residues. The O-linked sugars are attached to proteins through the oxygen atoms in serine, threonine, hydroxylysine, or hydroxylproline residues. The N-linked sugars are attached to proteins through the nitrogen atom in asparagine.
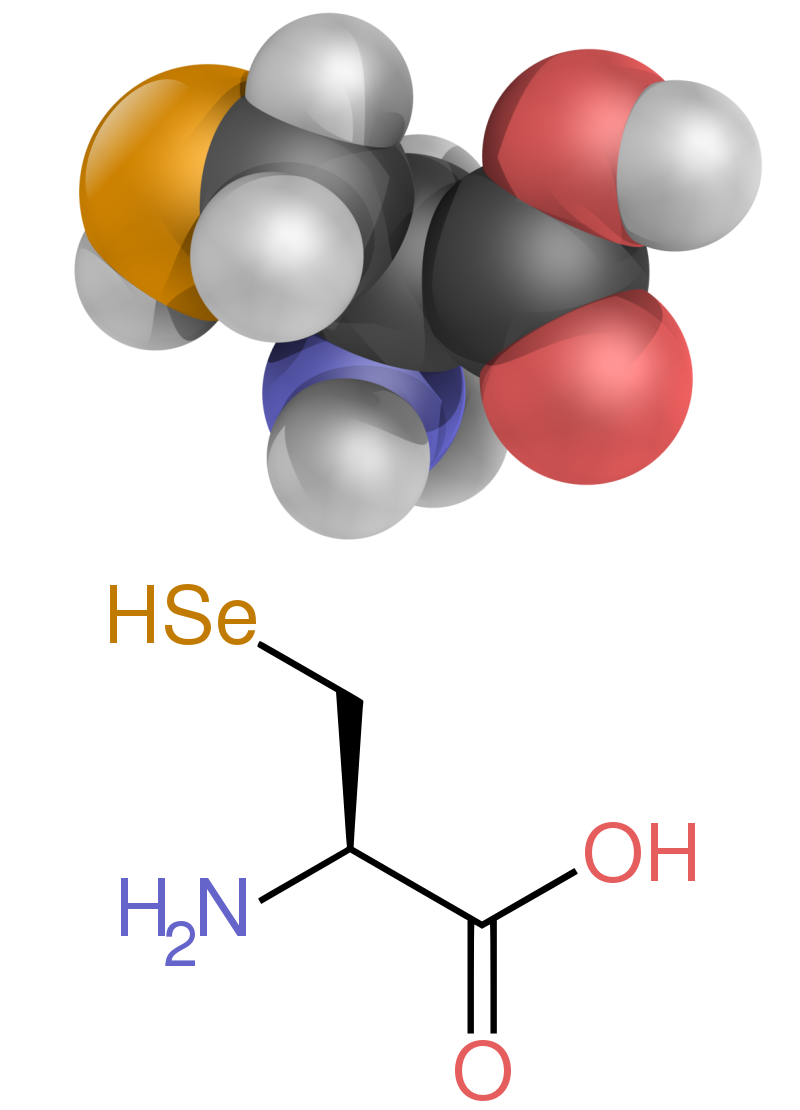
Finally, there is the case of selenocysteine. Although it is part of only a few known proteins, there is a sound scientific reason to consider this the 21st amino acid because it is in fact introduced during protein biosynthesis rather than created by a posttranslational modification. Selenocysteine is actually derived from the amino acid serine (in a very complicated fashion), and it contains selenium instead of the sulfur of cysteine.
The modern biochemist has a wide array of methods available for the separation and analysis of amino acids and proteins. These methods exploit the chemical differences of amino acids and in particular their ionization and solubility behaviour.
A typical determination of the amino acid composition of proteins involves three basic steps:
Hydrolysis is accomplished by treatment of a purified protein with a concentrated acid solution (6N HCl) at a very high temperature (usually 110 °C [230 °F]) for up to 70 hours. These conditions cleave the peptide bond between each and every amino acid residue.
The hydrolyzed protein sample is then separated into its constituent amino acids. Methods important for amino acid separations include ion exchange chromatography, gas chromatography, high-performance liquid chromatography, and most recently, capillary zone electrophoresis.
The sensitivity of the analysis of separated amino acids has been greatly improved by the use of fluorescent molecules that are attached to the amino acids, followed by their subsequent detection using fluorescence spectroscopy. For example, amino acids may be chemically “tagged” with a small fluorescent molecule (such as o-phthalaldehyde). These approaches routinely allow as little as a picomole (10−12 mole) of an amino acid to be detected. Most recently, this range of sensitivity has been extended to the attomole (10−18 mole) range.
The industrial production of amino acids is an important worldwide business. The first report of the commercial production of an amino acid was in 1908. It was then that the flavouring agent monosodium glutamate (MSG) was prepared from a type of large seaweed. This led to the commercial production of MSG, which is now produced using a bacterial fermentation process with starch and molasses as carbon sources. Glycine,cysteine, and D,L-alanine are also used as food additives, and mixtures of amino acids serve as flavour enhancers in the food industry. The amino acid balance of soy or corn protein for animal feed is significantly enhanced upon the addition of the nutritionally limiting amino acids methionine and lysine.
Monosodium glutamate (MSG)
Monosodium glutamate (MSG) crystals.
Amino acids are used therapeutically for nutritional and pharmaceutical purposes. For example, patients are often infused with amino acids to supply these nutrients before and after surgical procedures. Treatments with single amino acids are part of the medical approach to control certain disease states. Examples include L-dihydroxyphenylalanine (L-dopa) for Parkinson disease;glutamine and histidine to treat peptic ulcers; and arginine, citrulline, and ornithine to treat liver diseases.
Certain derivations of amino acids, especially of glutamate, are used as surfactants in mild soaps and shampoos. D-Phenylglycine and D-hydroxyphenylglycine are intermediates used for the chemical synthesis of β-lactam antibiotics (e.g., synthetic versions of penicillin). Aspartame is a sweetener prepared from the individual component amino acids aspartic acid and phenylalanine.
The question of why organisms on Earth consist of L-amino acids instead of D-amino acids is still an unresolved riddle. Some scientists have long suggested that a substantial fraction of the organic compounds that were the precursors to amino acids — and perhaps some amino acids themselves — on early Earth may have been derived from comet and meteorite impacts. One such organic-rich meteorite impact occurred on September 28, 1969, over Murchison, Victoria, Australia. This meteorite is suspected to be of cometary origin because of its high water content of 12 percent. Dozens of different amino acids have been identified within the Murchison meteorite, some of which are found on Earth. Some compounds identified in the meteorite, however, have no apparent terrestrial source. Most intriguing are the reports that amino acids in the Murchison meteorite exhibit an excess of L-amino acids. An extraterrestrial source for an L-amino acid excess in the solar system could predate the origin of life on Earth and thus explain the presence of a similar excess of L-amino acids on the prelife Earth.
In the form of proteins, amino acid residues form the second-largest component (water is the largest) of human muscles and other tissues. Beyond their role as residues in proteins, amino acids participate in a number of processes such as neurotransmitter transport and biosynthesis.
Nine proteinogenic amino acids are called “essential” for humans because they cannot be produced from other compounds by the human body and so must be taken in as food. Others may be conditionally essential for certain ages or medical conditions. Essential amino acids may also differ between species.
The first few amino acids were discovered in the early 19th century.In 1806, French chemists Louis-Nicolas Vauquelin and Pierre Jean Robiquet isolated a compound in asparagus that was subsequently named asparagine, the first amino acid to be discovered.Cystine was discovered in 1810, although its monomer, cysteine, remained undiscovered until 1884. Glycine and leucine were discovered in 1820. The last of the 20 common amino acids to be discovered was threonine in 1935 by William Cumming Rose, who also determined the essential amino acids and established the minimum daily requirements of all amino acids for optimal growth.
The unity of the chemical category was recognized by Wurtz in 1865, but he gave no particular name to it. First use of the term "amino acid" in the English language dates from 1898, while the German term, Aminosäure, was used earlier. Proteins were found to yield amino acids after enzymatic digestion or acid hydrolysis. In 1902, Emil Fischer and Franz Hofmeister independently proposed that proteins are formed from many amino acids, whereby bonds are formed between the amino group of one amino acid with the carboxyl group of another, resulting in a linear structure that Fischer termed "peptide".
In the structure shown at the top of the page, R represents a side chain specific to each amino acid. The carbon atom next to the carboxyl group (which is therefore numbered 2 in the carbon chain starting from that functional group) is called the α–carbon. Amino acids containing an amino group bonded directly to the alpha carbon are referred to as alpha amino acids. These include amino acids such as proline which contain secondary amines, which used to be often referred to as "imino acids".
The alpha amino acids are the most common form found in nature, but only when occurring in the L-isomer. The alpha carbon is a chiral carbon atom, with the exception of glycine which has two indistinguishable hydrogen atoms on the alpha carbon. Therefore, all alpha amino acids but glycine can exist in either of two enantiomers, called L or D amino acids (relative configuration), which are mirror images of each other (see also Chirality). While L-amino acids represent all of the amino acids found in proteins during translation in the ribosome, D-amino acids are found in some proteins produced by enzyme posttranslational modifications after translation and translocation to the endoplasmic reticulum, as in exotic sea-dwelling organisms such as cone snails. They are also abundant components of the peptidoglycancell walls of bacteria, and D-serine may act as a neurotransmitter in the brain. D-amino acids are used in racemic crystallography to create centrosymmetric crystals, which (depending on the protein) may allow for easier and more robust protein structure determination. The L and D convention for amino acid configuration refers not to the optical activity of the amino acid itself but rather to the optical activity of the isomer of glyceraldehyde from which that amino acid can, in theory, be synthesized (D-glyceraldehyde is dextrorotatory; L-glyceraldehyde is levorotatory). In alternative fashion, the (S) and (R) designators are used to indicate the absolute configuration. Almost all of the amino acids in proteins are (S) at the α carbon, with cysteine being (R) and glycine non-chiral. Cysteine has its side chain in the same geometric position as the other amino acids, but the R/S terminology is reversed because of the higher atomic number of sulfur compared to the carboxyl oxygen gives the side chain a higher priority by the Cahn–Ingold–Prelog rules, whereas the atoms in other side chains give them lower priority compared to the carboxyl group.
Amino acids are designated as α- when the nitrogen atom is attached to the carbon atom adjacent to the carboxyl group: in this case the compound contains the sub-structure N-C-CO2. Amino acids with the sub-structure N-C-C-CO2 are classified as β- amino acids. γ- amino acids contain the sub-structure N-C-C-C-CO2, and so on.
The phrase "branched-chain amino acids" or BCAA refers to the amino acids having aliphatic side chains that are linear; these are leucine,isoleucine, and valine.Proline is the only proteinogenic amino acid whose side-group links to the α-amino group and, thus, is also the only proteinogenic amino acid containing a secondary amine at this position. In chemical terms, proline is, therefore, an imino acid, since it lacks a primary amino group, although it is still classed as an amino acid in the current biochemical nomenclature, and may also be called an "N-alkylated alpha-amino acid".
In aqueous solution amino acids exist in two forms (as illustrated at the right), the molecular form and the zwitterion form in equilibrium with each other. The two forms co-exist over the pH range pK1 - 2 to pK2 + 2, which for glycine is pH 0-12. The ratio of the concentrations of the two isomers is independent of pH. The value of this ratio cannot be determined experimentally.
Because all amino acids contain amine and carboxylic acid functional groups, they are amphiprotic. At pH = pK1 (ca. 2.2) there will be equal concentration of the species NH3+CH(R)CO2H and NH3+CH(R)CO2- and at pH = pK2 (ca. 10) there will be equal concentration of the species NH3+CH(R)CO2- and NH2CH(R)CO2-. It follows that the neutral molecule and the zwitterion are effectively the only species present at biological pH.
It is generally assumed that the concentration of the zwitterion is much greater than the concentration of the neutral molecule on the basis of comparisons with the known pK values of amines and carboxylic acids.
An amino acid in its (1) molecular and (2) zwitterionic forms.
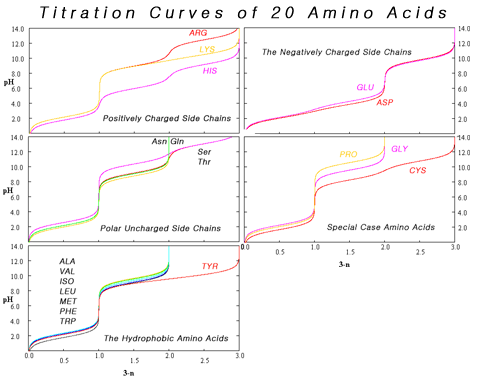
The variation in titration curves when the amino acids can be grouped by category. With the exception of tyrosine, using titration to distinguish among hydrophobic amino acids is problematic.
At pH values between the two pKa values, the zwitterion predominates, but coexists in dynamic equilibrium with small amounts of net negative and net positive ions. At the exact midpoint between the two pKa values, the trace amount of net negative and trace of net positive ions exactly balance, so that average net charge of all forms present is zero. This pH is known as the isoelectric point pI, so pI = ½(pKa1 + pKa2). The individual amino acids all have slightly different pKa values and therefore have different isoelectric points. For amino acids with charged side chains, the pKa of the side chain is involved. Thus for Asp or Glu with negative side chains, pI = ½(pKa1 + pKaR), where pKaR is the side chain pKa. Cysteine also has potentially negative side chain with pKaR = 8.14, so pI should be calculated as for Asp and Glu, even though the side chain is not significantly charged at physiological pH. For His, Lys, and Arg with positive side chains, pI = ½(pKaR + pKa2). Amino acids have zero mobility in electrophoresis at their isoelectric point, although this behaviour is more usually exploited for peptides and proteins than single amino acids. Zwitterions have minimum solubility at their isoelectric point, and some amino acids (in particular, with non-polar side chains) can be isolated by precipitation from water by adjusting the pH to the required isoelectric point.
Composite of titration curves of twenty proteinogenic amino acids grouped by side chain category.
.
A polypeptide is an unbranched chain of amino acids.
Amino acids are the structural units (monomers) that make up proteins. They join together to form short polymer chains called peptides or longer chains called either polypeptides or proteins.These polymers are linear and unbranched, with each amino acid within the chain attached to two neighboring amino acids. The process of making proteins encoded by DNA/RNA genetic material is called translationand involves the step-by-step addition of amino acids to a growing protein chain by a ribozyme that is called a ribosome. The order in which the amino acids are added is read through the genetic code from an mRNA template, which is an RNA copy of one of the organism’s genes.
Twenty-two amino acids are naturally incorporated into polypeptides and are called proteinogenic or natural amino acids. Of these, 20 are encoded by the universal genetic code. The remaining 2, selenocysteine and pyrrolysine, are incorporated into proteins by unique synthetic mechanisms. Selenocysteine is incorporated when the mRNA being translated includes a SECIS element, which causes the UGA codon to encode selenocysteine instead of a stop codon.Pyrrolysine is used by some methanogenicarchaea in enzymes that they use to produce methane. It is coded for with the codon UAG, which is normally a stop codon in other organisms. This UAG codon is followed by a PYLIS downstream sequence.
A polypeptide is an unbranched chain of amino acids.
Non-proteinogenic amino acids that are found in proteins are formed by post-translational modification, which is modification after translation during protein synthesis. These modifications are often essential for the function or regulation of a protein. For example, the carboxylation of glutamate allows for better binding of calcium cations, and collagen contains hydroxyproline, generated by hydroxylation of proline. Another example is the formation of hypusine in the translation initiation factorEIF5A, through modification of a lysine residue. Such modifications can also determine the localization of the protein, e.g., the addition of long hydrophobic groups can cause a protein to bind to a phospholipid membrane.
Some non-proteinogenic amino acids are not found in proteins. Examples include 2-aminoisobutyric acid and the neurotransmitter gamma-aminobutyric acid. Non-proteinogenic amino acids often occur as intermediates in the metabolic pathways for standard amino acids – for example, ornithine and citrulline occur in the urea cycle, part of amino acid catabolism (see below). A rare exception to the dominance of α-amino acids in biology is the β-amino acid beta alanine (3-aminopropanoic acid), which is used in plants and microorganisms in the synthesis of pantothenic acid (vitamin B5), a component of coenzyme A.
Although D-isomers are uncommon in live organisms, gramicidin is a polypeptide made up from mixture of D- and L-amino acids. Other compounds containing D-amino acids are tyrocidine and valinomycin. These compounds disrupt bacterial cell walls, particularly in Gram-positive bacteria. As of 2011, only 837 D-amino acids were found in the Swiss-Prot database out of a total of 187 million amino acids analysed.
The 20 amino acids that are encoded directly by the codons of the universal genetic code are called standard or canonical amino acids. A modified form of methionine (N-formylmethionine) is often incorporated in place of methionine as the initial amino acid of proteins in bacteria, mitochondria and chloroplasts. Other amino acids are called non-standard or non-canonical. Most of the non-standard amino acids are also non-proteinogenic (i.e. they cannot be incorporated into proteins during translation), but two of them are proteinogenic, as they can be incorporated translationally into proteins by exploiting information not encoded in the universal genetic code.
The two non-standard proteinogenic amino acids are selenocysteine (present in many non-eukaryotes as well as most eukaryotes, but not coded directly by DNA) and pyrrolysine (found only in some archaea and one bacterium). The incorporation of these non-standard amino acids is rare. For example, 25 human proteins include selenocysteine (Sec) in their primary structure, and the structurally characterized enzymes (selenoenzymes) employ Sec as the catalytic moiety in their active sites. Pyrrolysine and selenocysteine are encoded via variant codons. For example, selenocysteine is encoded by stop codon and SECIS element.
When taken up into the human body from the diet, the 20 standard amino acids either are used to synthesize proteins, other biomolecules, or are oxidized to urea and carbon dioxide as a source of energy. The oxidation pathway starts with the removal of the amino group by a transaminase; the amino group is then fed into the urea cycle. The other product of transamidation is a keto acid that enters the citric acid cycle.Glucogenic amino acids can also be converted into glucose, through gluconeogenesis. Of the 20 standard amino acids, nine (His,Ile,Leu,Lys,Met,Phe,Thr,Trp and Val) are called essential amino acids because the human body cannot synthesize them from other compounds at the level needed for normal growth, so they must be obtained from food. In addition, cysteine,tyrosine, and arginine are considered semiessential amino acids, and taurine a semiessential aminosulfonic acid in children. The metabolic pathways that synthesize these monomers are not fully developed. The amounts required also depend on the age and health of the individual, so it is hard to make general statements about the dietary requirement for some amino acids. Dietary exposure to the non-standard amino acid BMAA has been linked to human neurodegenerative diseases, including ALS.
Some non-standard amino acids are used as defenses against herbivores in plants. For example, canavanine is an analogue of arginine that is found in many legumes, and in particularly large amounts in Canavalia gladiata (sword bean). This amino acid protects the plants from predators such as insects and can cause illness in people if some types of legumes are eaten without processing. The non-protein amino acid mimosine is found in other species of legume, in particular Leucaena leucocephala. This compound is an analogue of tyrosine and can poison animals that graze on these plants.
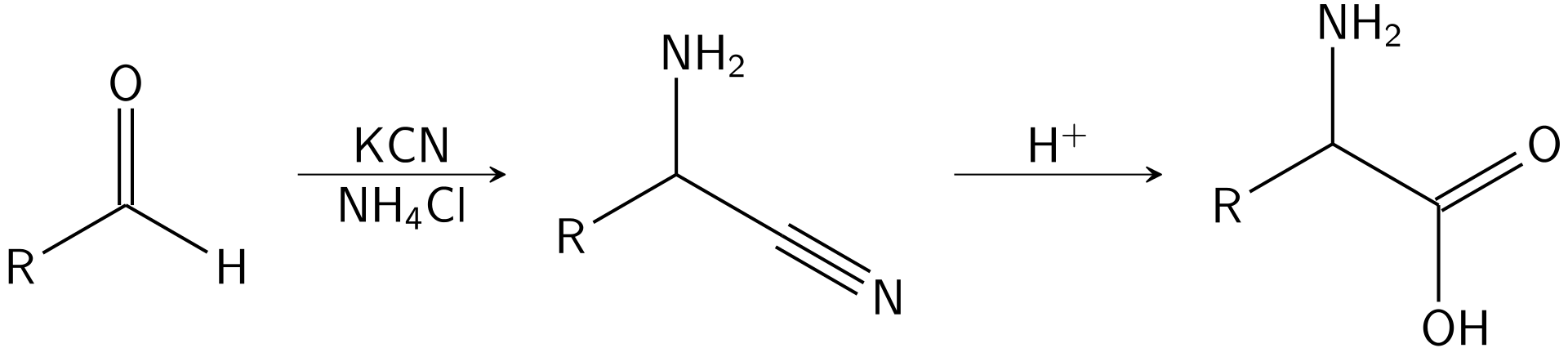
The commercial production of amino acids usually relies on mutant bacteria that overproduce individual amino acids using glucose as a carbon source. Some amino acids are produced by enzymatic conversions of synthetic intermediates. 2-Aminothiazoline-4-carboxylic acid is an intermediate in one industrial synthesis of L-cysteine for example. Aspartic acid is produced by the addition of ammonia to fumarate using a lyase.
In plants, nitrogen is first assimilated into organic compounds in the form of glutamate, formed from alpha-ketoglutarate and ammonia in the mitochondrion. For other amino acids, plants use transaminases to move the amino group from glutamate to another alpha-keto acids. For example, aspartate aminotransferase converts glutamate and oxaloacetate to alpha-ketoglutarate and aspartate. Other organisms use transaminases for amino acid synthesis, too.
Amino acids undergo the reactions expected of the constituent functional groups. The types of these reactions are determined by the groups on these side chains and are, therefore, different between the various types of amino acid.
As both the amine and carboxylic acid groups of amino acids can react to form amide bonds, one amino acid molecule can react with another and become joined through an amide linkage. This polymerization of amino acids is what creates proteins. This condensation reaction yields the newly formed peptide bond and a molecule of water. In cells, this reaction does not occur directly; instead, the amino acid is first activated by attachment to a transfer RNA molecule through an ester bond. This aminoacyl-tRNA is produced in an ATP-dependent reaction carried out by an aminoacyl tRNA synthetase. This aminoacyl-tRNA is then a substrate for the ribosome, which catalyzes the attack of the amino group of the elongating protein chain on the ester bond. As a result of this mechanism, all proteins made by ribosomes are synthesized starting at their N-terminus and moving toward their C-terminus.
However, not all peptide bonds are formed in this way. In a few cases, peptides are synthesized by specific enzymes. For example, the tripeptide glutathione is an essential part of the defenses of cells against oxidative stress. This peptide is synthesized in two steps from free amino acids. In the first step, gamma-glutamylcysteine synthetase condenses cysteine and glutamic acid through a peptide bond formed between the side chain carboxyl of the glutamate (the gamma carbon of this side chain) and the amino group of the cysteine. This dipeptide is then condensed with glycine by glutathione synthetase to form glutathione.
In chemistry, peptides are synthesized by a variety of reactions. One of the most-used in solid-phase peptide synthesis uses the aromatic oxime derivatives of amino acids as activated units. These are added in sequence onto the growing peptide chain, which is attached to a solid resin support. The ability to easily synthesize vast numbers of different peptides by varying the types and order of amino acids (using combinatorial chemistry) has made peptide synthesis particularly important in creating libraries of peptides for use in drug discovery through high-throughput screening.
The combination of functional groups allow amino acids to be effective polydentate ligands for metal-amino acid chelates. The multiple side chains of amino acids can also undergo chemical reactions.
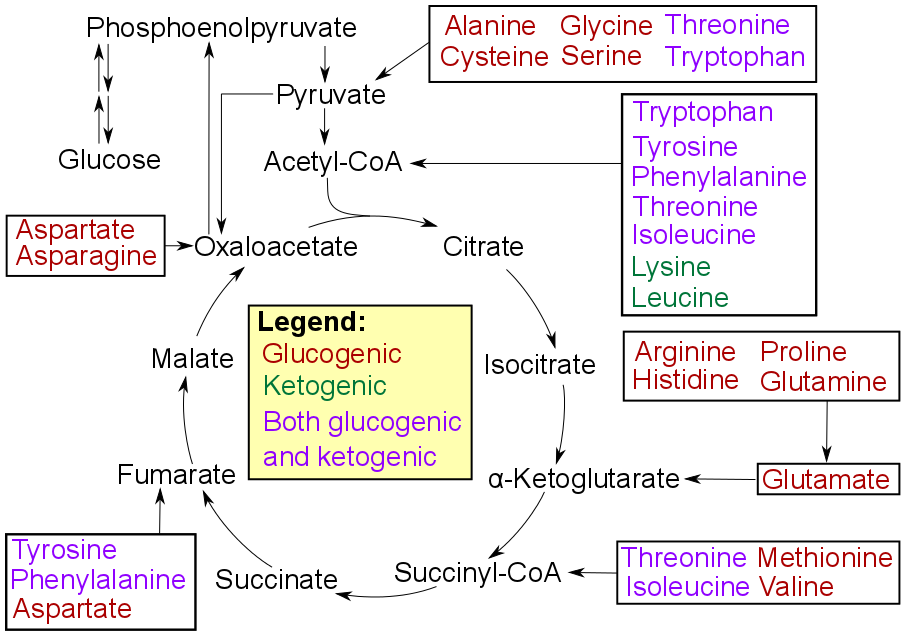
Amino acids must first pass out of organelles and cells into blood circulation via amino acid transporters, since the amine and carboxylic acid groups are typically ionized. Degradation of an amino acid, occurring in the liver and kidneys, often involves deamination by moving its amino group to alpha-ketoglutarate, forming glutamate. This process involves transaminases, often the same as those used in amination during synthesis. In many vertebrates, the amino group is then removed through the urea cycle and is excreted in the form of urea. However, amino acid degradation can produce uric acid or ammonia instead. For example, serine dehydratase converts serine to pyruvate and ammonia. After removal of one or more amino groups, the remainder of the molecule can sometimes be used to synthesize new amino acids, or it can be used for energy by entering glycolysis or the citric acid cycle, as detailed in image at right.
Catabolism of proteinogenic amino acids. Amino acids can be classified according to the properties of their main products as either of the following: * Glucogenic, with the products having the ability to form glucose by gluconeogenesis * Ketogenic, with the products not having the ability to form glucose. These products may still be used for ketogenesis or lipid synthesis. * Amino acids catabolized into both glucogenic and ketogenic products..



.png)