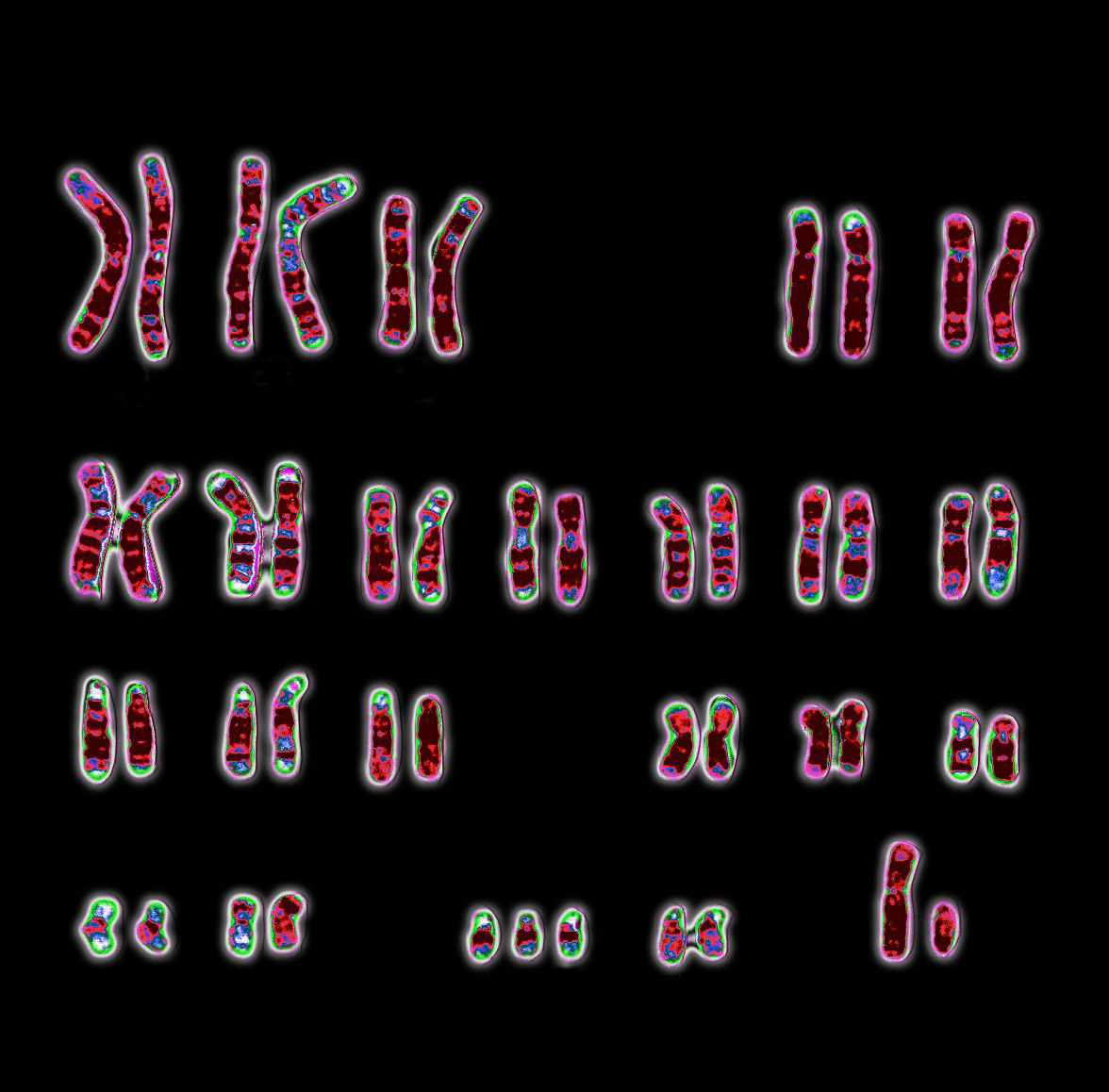Mutasyon bir örgenlik ya da virus genomunun ya da kromozom-dışı DNAnın nükleotid dizisinde kalıtlanabilir bir değişimdir.
Mutasyon DNA eşlenimi, viral eşlenim, mitosis ve meiosis sırasında oluşani hatalardan ya da başka DNA hasarlarından doğar.
Mutasyonlar bir örgenliğin fenotipinde belirgin değişimler yaratabilir ya da yaratmayabilir.
Mutasyon evrimde, immün dizgenin gelişiminde, kanserde rol oynar.
Mutasyon genlerde hiçbir etki yapmayabilir, bir gen ürününü değiştirebilir, ya da genin işlevini çeşitli düzeylerde ya da tam olarak engelleyebilir.
MUTASYONUN NEDENLERİ
Kendiliğinden mutasyonlar (moleküler bozunum).
Doğallıkla olan DNA hasarının atlanmasından kaynaklanan hatalı eşlenim.
DNA onarımı sırasında ortaya çıkan hatalar.
Mutajenlerin yol açtığı irkilim mutasyonları.
GENLER
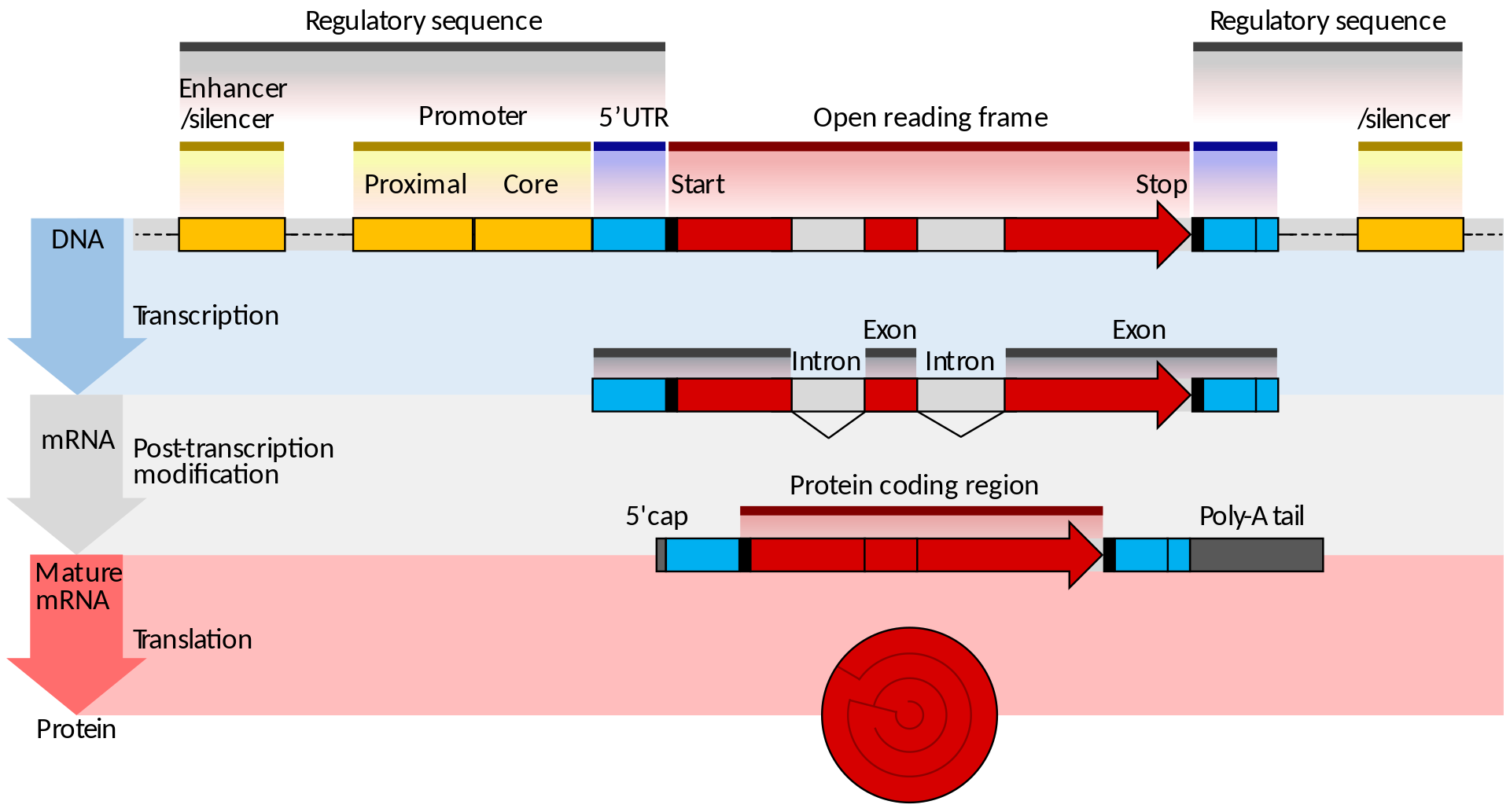
Genler DNAnın işlevsel birimleridir.
Genin düzenleyici bölgesi eşyazım sürecinin açık ya da kapalı olmasını belirler.
Genin kodlayıcı bölgesi genellikle bir protein olan işlevsel molekülün yapısını belirleyen genetik kodu taşır.
Bir genin mutasyona uğramış biçimi olan “mutantallel”de değişim ya düzenleyici bölgede ya da kodlayıcı bölgede yer alır.
Bir protein 20 ortak amino asidin belirli bir sayı ve belirli bir diziliş içinde yapılanan bir polipeptid zinciridir.
Her bir amino asit olanaklı dört baz çiftinin üçünden oluşan benzersiz bir kodon tarafından kodlanır (DNAda baz çiftleri: A–T, T–A, G–C, ve C–G; ve dört azotlu baz: adenin, thimin, guanin ve sitozin).
DNA dizisini değiştiren bir mutason amino asit dizisini de değiştirir ve proteinin işlevini indirger ya da bozar.
Genin düzenleyici bölgesindeki bir değişim protein üretiminin zamanlamasını ve olnağını zararlı olarak etkileyebilir ve hücrede işle bozukluğuna götürebilir.
Sessiz mutasyonlar işlevsel düzlemde açık bir etki göstermeyen değişimlerdir.
MUTASYON TİPLERİ NOKTA MUTASYON
Genlerdeki değişimlere nokta mutasyonlar denir.
Nokta mutasyonun en yalın tipi tekil baz-çiftlerinin yer değiştirmesidir ve sonuç proteinde değişmiş bir amino asit dizisi ya da erken bir konuma giren bir dur kodonudur.
Tekil baz-çiftlerini araya sokan ya da silen değişimler de protein yapısını bozar.
Baz-çiftlerinin daha karmaşık bileşimleri de olabilir.
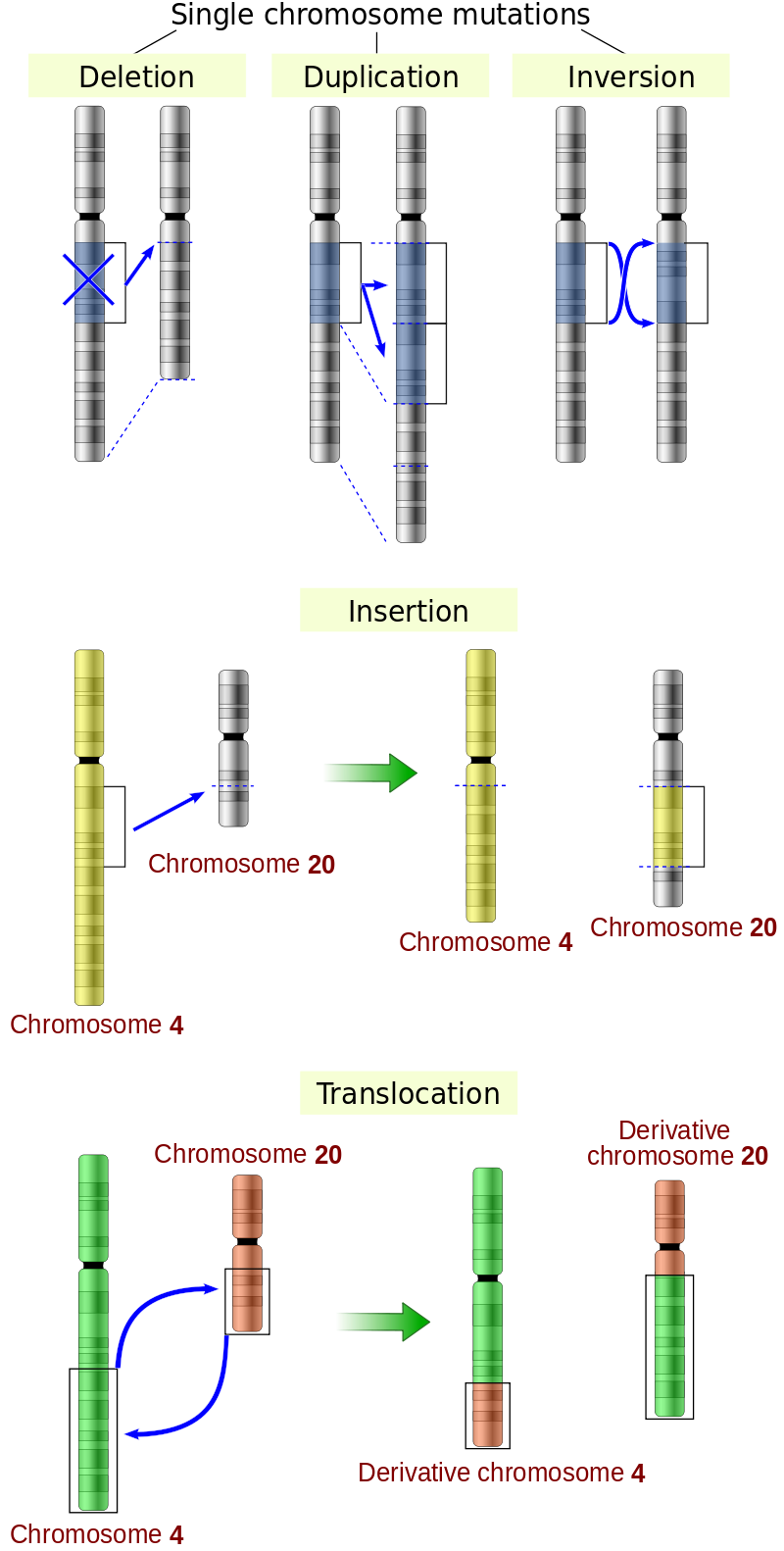
KROMOZOMAL MUTASYON
Birden daha çok gene yayılan mutasyonlara kromozomal mutasyonlar denir.
Kromozomal mutasyonlar büyük ölçek silinme, ikileme, evrilme ve yerleşim bozulmalarından oluşur.
Kromozomlarda yer alan mutasyonlar bütün DNA molekülünün yapı, işlev ve kalıtımını etkiler.
Silinme ve ikilemeler sık sık büyük ölçek anomalilerde sonuçlanır.
Evrilme ve yerleşim bozuklukları bir gen içerisinde kopma olmadıkça işlevsel bozukluğa yol açmaz.
Bütün kromozomların yitim ya da kazanımı (aneuploidy) ciddi anomalilerde (örneğin fazla bir kromozom 21’in yol açtığı Down sendromu) ya da ölümde sonuçlanabilir.
Bütün kromozom kümelerinin kazanımı (iki yerine üç, dört ya da daha çok kromozom kümesinin bulunuşu, polyploidy) yeni türlerin evriminde rol oynar.
🛑 MUTASYON VE EVRİM — DİZİ BENZEŞİMİ
MUTASYON VE EVRİM
Mutasyonlar genellikle genetik yeniden-bileşim yoluyla olmak üzere DNAnın büyük kesimlerinin eşlenmesini içerebilir.
Bu yeniden-bileşimler ve eşlenimler yeni genlerin evrimine yol açar.
TERİMLER
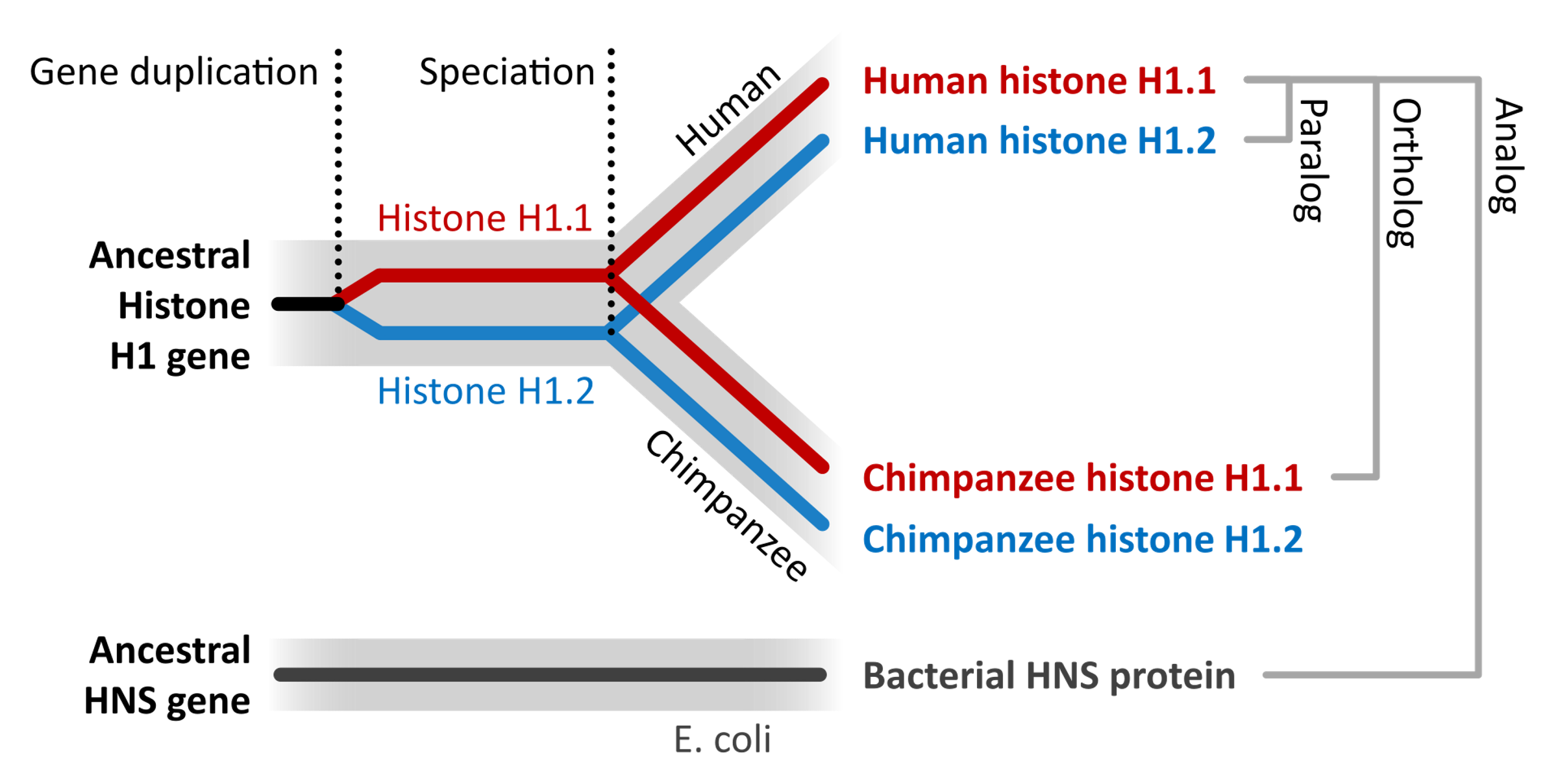
homolog: Homolog genler ayrı taksonlara ait iki birey arasında ortak ata nedeniyle benzer olan genlerdir.
homoeolog: Homoeolg kromozomlar ya da kromozom kesimleri türler-arası hibridizasyon sonucunda biraraya gelerek hibrid bir genom oluştururlar.
ohnolog: Ohnolgo genler 2R bütün-genom eşlenimi süreci yoluyla ortaya çıkan paralog genlerdir (ad ‘Susumu Ohno’dan gelir).
ortholog: Ortholog genler değişik türlerde ortak bir ata geninden türleşme yoluyla evrimlenen genlerdir; genel olarak, ortlologlar evrim süreci boyunca aynı işlevi sürdürürler. (L)
paralog: Paralog genler karşılaştırılan türlerin son ortak atasındaki eşlenim olayları yoluyla ilişkili genlerdir.
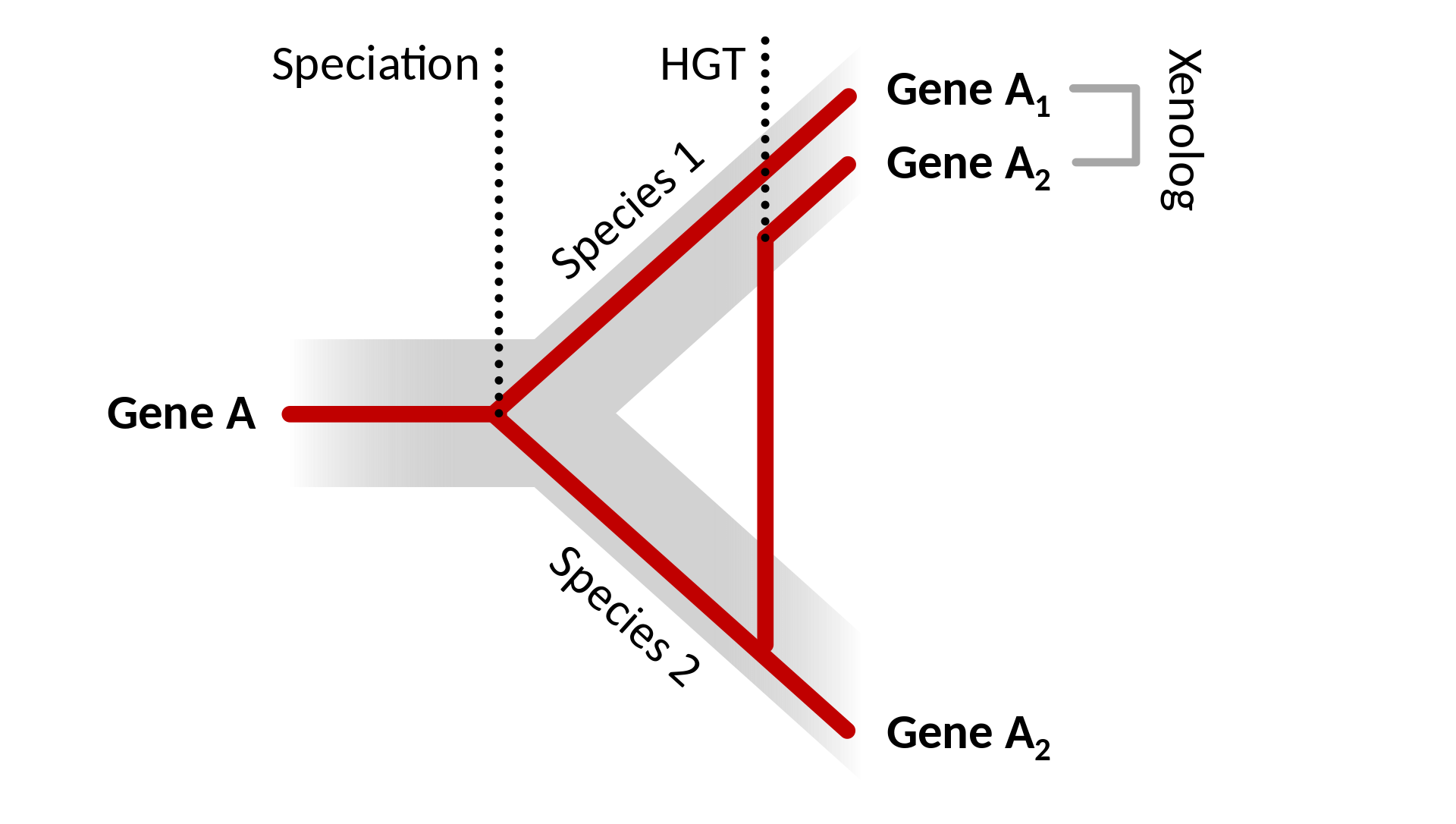
xenolog: Xenologlar iki örgenlik arasındaki yatay gen aktarımından ortaya çıkan genlerdir.
DİZİ BENZEŞİMİ
Dizi benzeşmesi evrim sürecinde DNA, RNA ya da protein dizileri arasında ortak ataların terimlerinde tanımlanan biolojik benzeşikliktir.
İki DNA kesimi ya (1) türleşme olayı (orthologos), ya (2) eşlenim olayı (paralogos) ya da (3) yatay gen aktarımı olayı (xenologos) nedeniyle ortak atadan geliyor olabilir.
DNA, RNA ya da proteinler arasındaki benzerlik tipik olarak nükleotid ya da amino asit dizilerinin benzerliğinden çıkarsanır.
Nükleotid ya da amino asit dizileri arasındaki benzerlik dizilerin ortak bir ata dizinin uğradığı evrimsel değişimler yoluyla ilişkili olduğu konusunda güçlü bir kanıt sunabilir.
Türsel evrim süreci (gri) içerisinde yer alan gen evrim süreci (kırmızı ve mavi). Üst: Bir ata gen eşlenimi iki paralog üretir (histon H1.1 ve 1.2). Bir türleşme olayı iki yeni kuşak türde (insan ve şempanze) orthologlar üretir.
Alt: Ayrı bir türde (E. coli) bir genin benzer bir işlevi (histon-benzeri nükeoid-yapıcı protein) vardır, ama ayrı bir evrimsel köken taşır ve böylece bir analogtur. (W)
Üst: Bir ata geni iki paralog (A ve B genleri) üretmek üzere eşlenir. Bir türleşme olayı iki yeni kuşak türde orthologlar üretir. Alt: Ayrı bir türde ilişkisiz bir genin benzer bir işlevi vardır (C geni), ama
ayrı bir evrimsel süreçten gelir ve böylece bir analogtur.
Bütün bir genom eşlenim olayı her bir genin iki ohnolog eşlemi ile bir genom üretir.
Bir türleşme olayı iki yeni kuşak türde bir genin ortholglarını üretir. Bir türden bir başkasına bir yatay gen aktarımı olayı genin genomuna bir xenolog ekler.
Bir türleşme olayı iki yeni kuşak türde bir genin orthologlarını üretir. Bu türlerin sonraki hibridleşmesi her iki türden her bir genin bir homoeolog eşlemi ile bir hibrid genom üretir.
First-ever look at a true 3D model of DNA replication/repair. This project was done in Maya under the direction of a pre-med student and instructor. Hopefully, this simple illustration helps give you a better idea of how our bodies work!
📹 The Effects of Mutation (Secrets of the Cell with Michael Behe, Ep. 4) (VİDEO)
📹 The Effects of Mutation (Secrets of the Cell with Michael Behe, Ep. 4) (LINK)
Biochemist and bestselling author Michael Behe explores the impact of mutations on evolution in Episode 4 of his series "Secrets of the Cell with Michael Behe." Do random mutations and natural selection provide an explanation for the development of new organisms and features in the history of life?
Mutation, an alteration in the genetic material (the genome)of a cell of a living organism or of a virus that is more or less permanent and that can be transmitted to the cell’s or the virus’s descendants. (The genomes of organisms are all composed of DNA, whereas viral genomes can be of DNA or RNA; see heredity: The physical basis of heredity.) Mutation in the DNA of a body cell of a multicellular organism( somatic mutation) may be transmitted to descendant cells by DNA replication and hence result in a sector or patch of cells having abnormal function, an example being cancer. Mutations in egg or sperm cells ( germinal mutations) may result in an individual offspring all of whose cells carry the mutation, which often confers some serious malfunction, as in the case of a human genetic disease such as cystic fibrosis. Mutations result either from accidents during the normal chemical transactions of DNA, often during replication, or from exposure to high-energy electromagnetic radiation (e.g., ultraviolet light or X-rays) or particle radiation or to highly reactive chemicals in the environment. Because mutations are random changes, they are expected to be mostly deleterious, but some may be beneficial in certain environments. In general, mutation is the main source of genetic variation, which is the raw material {?} for evolution by natural selection{?}.
Point mutation
The effect of base substitutions, or point mutations, on the messenger-RNA codon AUA, which codes for the amino acid isoleucine. Substitutions (red letters) at the first, second, or third position in the codon can result in nine new codons corresponding to six different amino acids in addition to isoleucine itself. The chemical properties of some of these amino acids are quite different from those of isoleucine. Replacement of one amino acid in a protein by another can seriously affect the protein's biological function.
📹 Mutation
The genome is composed of one to several long molecules of DNA, and mutation can occur potentially anywhere on these molecules at any time. The most serious changes take place in the functional units of DNA, the genes.A mutated form of a gene is called a mutantallele.A gene is typically composed of a regulatory region, which is responsible for turning the gene’s transcription on and off at the appropriate times during development, and a coding region, which carries the genetic code for the structure of a functional molecule, generally a protein.A protein is a chain of usually several hundred amino acids. Cells make 20 common amino acids, and it is the unique number and sequence of these that give a protein its specific function. Each amino acid is encoded by a unique sequence, or codon, of three of the four possible base pairs in the DNA (A–T, T–A, G–C, and C–G, the individual letters referring to the four nitrogenous bases adenine,thymine,guanine, and cytosine). Hence, a mutation that changes DNA sequence can change amino acid sequence and in this way potentially reduce or inactivate a protein’s function. A change in the DNA sequence of a gene’s regulatory region can adversely affect the timing and availability of the gene’s protein and also lead to serious cellular malfunction. On the other hand, many mutations are silent, showing no obvious effect at the functional level. Some silent mutations are in the DNA between genes, or they are of a type that results in no significant amino acid changes.
Mutations are of several types. Changes within genes are called point mutations. The simplest kinds are changes to single base pairs, called base-pair substitutions. Many of these substitute an incorrect amino acid in the corresponding position in the encoded protein, and of these a large proportion result in altered protein function. Some base-pair substitutions produce a stop codon. Normally, when a stop codon occurs at the end of a gene, it stops protein synthesis, but, when it occurs in an abnormal position, it can result in a truncated and nonfunctional protein. Another type of simple change, the deletion or insertion of single base pairs, generally has a profound effect on the protein because the protein’s synthesis, which is carried out by the reading of triplet codons in a linear fashion from one end of the gene to the other, is thrown off. This change leads to a frameshift in reading the gene such that all amino acids are incorrect from the mutation onward. More-complex combinations of base substitutions, insertions, and deletions can also be observed in some mutant genes.
Mutations that span more than one gene are called chromosomal mutations because they affect the structure, function, and inheritance of whole DNA molecules (microscopically visible in a coiled state as chromosomes). Often these chromosome mutations result from one or more coincident breaks in the DNA molecules of the genome (possibly from exposure to energetic radiation), followed in some cases by faulty rejoining. Some outcomes are large-scale deletions, duplications, inversions, and translocations. In a diploid species (a species, such as human beings, that has a double set of chromosomes in the nucleus of each cell), deletions and duplications alter gene balance and often result in abnormality. Inversions and translocations involve no loss or gain and are functionally normal unless a break occurs within a gene. However, at meiosis (the specialized nuclear divisions that take place during the production of gametes — i.e., eggs and sperm), faulty pairing of an inverted or translocated chromosome set with a normal set can result in gametes and hence progeny with duplications and deletions.
Loss or gain of whole chromosomes results in a condition called aneuploidy. One familiar result of aneuploidy is Down syndrome, a chromosomal disorder in which humans are born with an extra chromosome 21 (and hence bear three copies of that chromosome instead of the usual two). Another type of chromosome mutation is the gain or loss of whole chromosome sets. Gain of sets results in polyploidy—that is, the presence of three, four, or more chromosome sets instead of the usual two. Polyploidy has been a significant force in the evolution of new species of plants and animals. (See alsoevolution: Polyploidy.)
Karyotype; Down syndrome
A karyotype of a human male with Down syndrome, showing a full chromosome complement plus an extra chromosome 21.
Most genomes contain mobile DNA elements that move from one location to another. The movement of these elements can cause mutation, either because the element arrives in some crucial location, such as within a gene, or because it promotes large-scale chromosome mutations via recombination between pairs of mobile elements in different locations.
At the level of whole populations of organisms, mutation can be viewed as a constantly dripping faucet introducing mutant alleles into the population, a concept described as mutational pressure. The rate of mutation differs for different genes and organisms. In RNA viruses, such as the human immunodeficiency virus (HIV; seeAIDS), replication of the genome takes place within the host cell using a mechanism that is prone to error. Hence, mutation rates in such viruses are high. In general, however, the fate of individual mutant alleles is never certain. Most are eliminated by chance. In some cases a mutant allele can increase in frequency by chance, and then individuals expressing the allele can be subject to selection, either positive or negative. Hence, for any one gene the frequency of a mutant allele in a population is determined by a combination of mutational pressure, selection, and chance.
Somatic mutation, genetic alteration acquired by a cell that can be passed to the progeny of the mutated cell in the course of cell division. Somatic mutations differ from germ line mutations, which are inherited genetic alterations that occur in the germ cells (i.e., sperm and eggs). Somatic mutations are frequently caused by environmental factors, such as exposure to ultraviolet radiation or to certain chemicals.
Somatic mutations may occur in any cell division from the first cleavage of the fertilized egg to the cell divisions that replace cells in a senile individual. The mutation affects all cells descended from the mutated cell. A major part of an organism, such as the branch of a tree or a complete tissue layer of an animal, may carry the mutation; it may or may not be expressed visibly. Somatic mutations can give rise to various diseases, including cancer.
Germinal mutation (genetics)
Germinal mutation (genetics)
Germinal mutation (genetics) (B) Germinal mutation, alteration in the genetic constitution of the reproductive cells, occurring in the cell divisions that result in sperm and eggs. Germinal mutations can be caused by radiation or chemical mutagens and may affect a single gene or an entire chromosome. A germinal mutation affects the progeny of the individual in whose reproductive cells the mutation arose and subsequent generations of that progeny. Unlike somatic mutations, which occur in the body cells and are not passed on to later generations, germinal mutations are important sources of genetic variation in natural populations that lead to evolutionary change through natural selection.
Mutations may or may not produce discernible changes in the observable characteristics (phenotype) of an organism. Mutations play a part in both normal and abnormal biological processes including:evolution,cancer, and the development of the immune system,including junctional diversity. Mutation is the ultimate source of all genetic variation, providing the raw material on which evolutionary forces such as natural selection can act.
Mutation can result in many different types of change in sequences. Mutations in genescan have no effect, alter the product of a gene, or prevent the gene from functioning properly or completely. Mutations can also occur in nongenic regions. A 2007 study on genetic variations between different species of Drosophila suggested that, if a mutation changes a protein produced by a gene, the result is likely to be harmful, with an estimated 70% of amino acidpolymorphisms that have damaging effects, and the remainder being either neutral or marginally beneficial. Due to the damaging effects that mutations can have on genes, organisms have mechanisms such as DNA repair to prevent or correct mutations by reverting the mutated sequence back to its original state.
Mutations can involve the duplication of large sections of DNA, usually through genetic recombination. These duplications are a major source of raw material for evolving new genes, with tens to hundreds of genes duplicated in animal genomes every million years. Most genes belong to larger gene families of shared ancestry, detectable by their sequence homology. Novel genes are produced by several methods, commonly through the duplication and mutation of an ancestral gene, or by recombining parts of different genes to form new combinations with new functions.
Here, protein domains act as modules, each with a particular and independent function, that can be mixed together to produce genes encoding new proteins with novel properties. For example, the human eye uses four genes to make structures that sense light: three for cone cell or color vision and one for rod cell or night vision; all four arose from a single ancestral gene. Another advantage of duplicating a gene (or even an entire genome) is that this increases engineering redundancy; this allows one gene in the pair to acquire a new function while the other copy performs the original function. Other types of mutation occasionally create new genes from previously noncoding DNA.
Changes in chromosome number may involve even larger mutations, where segments of the DNA within chromosomes break and then rearrange. For example, in the Homininae, two chromosomes fused to produce human chromosome 2; this fusion did not occur in the lineage of the other apes, and they retain these separate chromosomes. In evolution, the most important role of such chromosomal rearrangements may be to accelerate the divergence of a population into new species by making populations less likely to interbreed, thereby preserving genetic differences between these populations.
Sequences of DNA that can move about the genome, such as transposons, make up a major fraction of the genetic material of plants and animals, and may have been important in the evolution of genomes. For example, more than a million copies of the Alu sequence are present in the human genome, and these sequences have now been recruited to perform functions such as regulating gene expression. Another effect of these mobile DNA sequences is that when they move within a genome, they can mutate or delete existing genes and thereby produce genetic diversity.
Nonlethal mutations accumulate within the gene pool and increase the amount of genetic variation. The abundance of some genetic changes within the gene pool can be reduced by natural selection, while other "more favorable" mutations may accumulate and result in adaptive changes.
For example, a butterfly may produce offspring with new mutations. The majority of these mutations will have no effect; but one might change the color of one of the butterfly's offspring, making it harder (or easier) for predators to see. If this color change is advantageous, the chances of this butterfly's surviving and producing its own offspring are a little better, and over time the number of butterflies with this mutation may form a larger percentage of the population.
Neutral mutations are defined as mutations whose effects do not influence the fitness of an individual. These can increase in frequency over time due to genetic drift. It is believed that the overwhelming majority of mutations have no significant effect on an organism's fitness. Also, DNA repair mechanisms are able to mend most changes before they become permanent mutations, and many organisms have mechanisms for eliminating otherwise-permanently mutated somatic cells.
Beneficial mutations can improve reproductive success.
Four classes of mutations are (1) spontaneous mutations (molecular decay), (2) mutations due to error-prone replication bypass of naturally occurring DNA damage (also called error-prone translesion synthesis), (3) errors introduced during DNA repair, and (4) induced mutations caused by mutagens. Scientists may also deliberately introduce mutant sequences through DNA manipulation for the sake of scientific experimentation.
One 2017 study claimed that 66% of cancer-causing mutations are random, 29% are due to the environment (the studied population spanned 69 countries), and 5% are inherited.
Humans on average pass 60 new mutations to their children but fathers pass more mutations depending on their age with every year adding two new mutations to a child.
Spontaneous mutations occur with non-zero probability even given a healthy, uncontaminated cell. Naturally occurring oxidative DNA damage is estimated to occur 10,000 times per cell per day in humans and 100,000 times per cell per day in rats. Spontaneous mutations can be characterized by the specific change:
Tautomerism – A base is changed by the repositioning of a hydrogen atom, altering the hydrogen bonding pattern of that base, resulting in incorrect base pairing during replication.
Deamination – Hydrolysis changes a normal base to an atypical base containing a keto group in place of the original amine group. Examples include C → U and A → HX (hypoxanthine), which can be corrected by DNA repair mechanisms; and 5MeC (5-methylcytosine) → T, which is less likely to be detected as a mutation because thymine is a normal DNA base.
Slipped strand mispairing – Denaturation of the new strand from the template during replication, followed by renaturation in a different spot ("slipping"). This can lead to insertions or deletions.
Error-prone replication bypass (W)
There is increasing evidence that the majority of spontaneously arising mutations are due to error-prone replication (translesion synthesis) past DNA damage in the template strand. In mice, the majority of mutations are caused by translesion synthesis. Likewise, in yeast, Kunz et al. found that more than 60% of the spontaneous single base pair substitutions and deletions were caused by translesion synthesis.
Although naturally occurring double-strand breaks occur at a relatively low frequency in DNA, their repair often causes mutation. Non-homologous end joining (NHEJ) is a major pathway for repairing double-strand breaks. NHEJ involves removal of a few nucleotides to allow somewhat inaccurate alignment of the two ends for rejoining followed by addition of nucleotides to fill in gaps. As a consequence, NHEJ often introduces mutations.
Alkylating agents (e.g., N-ethyl-N-nitrosourea (ENU). These agents can mutate both replicating and non-replicating DNA. In contrast, a base analog can mutate the DNA only when the analog is incorporated in replicating the DNA. Each of these classes of chemical mutagens has certain effects that then lead to transitions,transversions, or deletions.
Nitrous acid converts amine groups on A and C to diazo groups, altering their hydrogen bonding patterns, which leads to incorrect base pairing during replication.
Whereas in former times mutations were assumed to occur by chance, or induced by mutagens, molecular mechanisms of mutation have been discovered in bacteria and across the tree of life. As S. Rosenberg states, "These mechanisms reveal a picture of highly regulated mutagenesis, up-regulated temporally by stress responses and activated when cells/organisms are maladapted to their environments—when stressed — potentially accelerating adaptation." Since they are self-induced mutagenic mechanisms that increase the adaptation rate of organisms, they have some times been named as adaptive mutagenesis mechanisms, and include the SOS response in bacteria, ectopic intrachromosomal recombination and other chromosomal events such as duplications.
By effect on structure (W)
The sequence of a gene can be altered in a number of ways. Gene mutations have varying effects on health depending on where they occur and whether they alter the function of essential proteins. Mutations in the structure of genes can be classified into several types.
Large-scale mutations in chromosomal structure include:
Amplifications (or gene duplications) or repetition of a chromosomal segment or presence of extra piece of a chromosome broken piece of a chromosome may become attached to a homologous or non-homologous chromosome so that some of the genes are present in more than two doses leading to multiple copies of all chromosomal regions, increasing the dosage of the genes located within them.
Deletions of large chromosomal regions, leading to loss of the genes within those regions.
Mutations whose effect is to juxtapose previously separate pieces of DNA, potentially bringing together separate genes to form functionally distinct fusion genes (e.g., bcr-abl).
Large scale changes to the structure of chromosomes called chromosomal rearrangement that can lead to a decrease of fitness but also to speciation in isolated, inbred populations. These include:
Interstitial deletions: an intra-chromosomal deletion that removes a segment of DNA from a single chromosome, thereby apposing previously distant genes. For example, cells isolated from a human astrocytoma, a type of brain tumor, were found to have a chromosomal deletion removing sequences between the Fused in Glioblastoma (FIG) gene and the receptor tyrosine kinase (ROS), producing a fusion protein (FIG-ROS). The abnormal FIG-ROS fusion protein has constitutively active kinase activity that causes oncogenic transformation (a transformation from normal cells to cancer cells).
Loss of heterozygosity: loss of one allele, either by a deletion or a genetic recombination event, in an organism that previously had two different alleles.
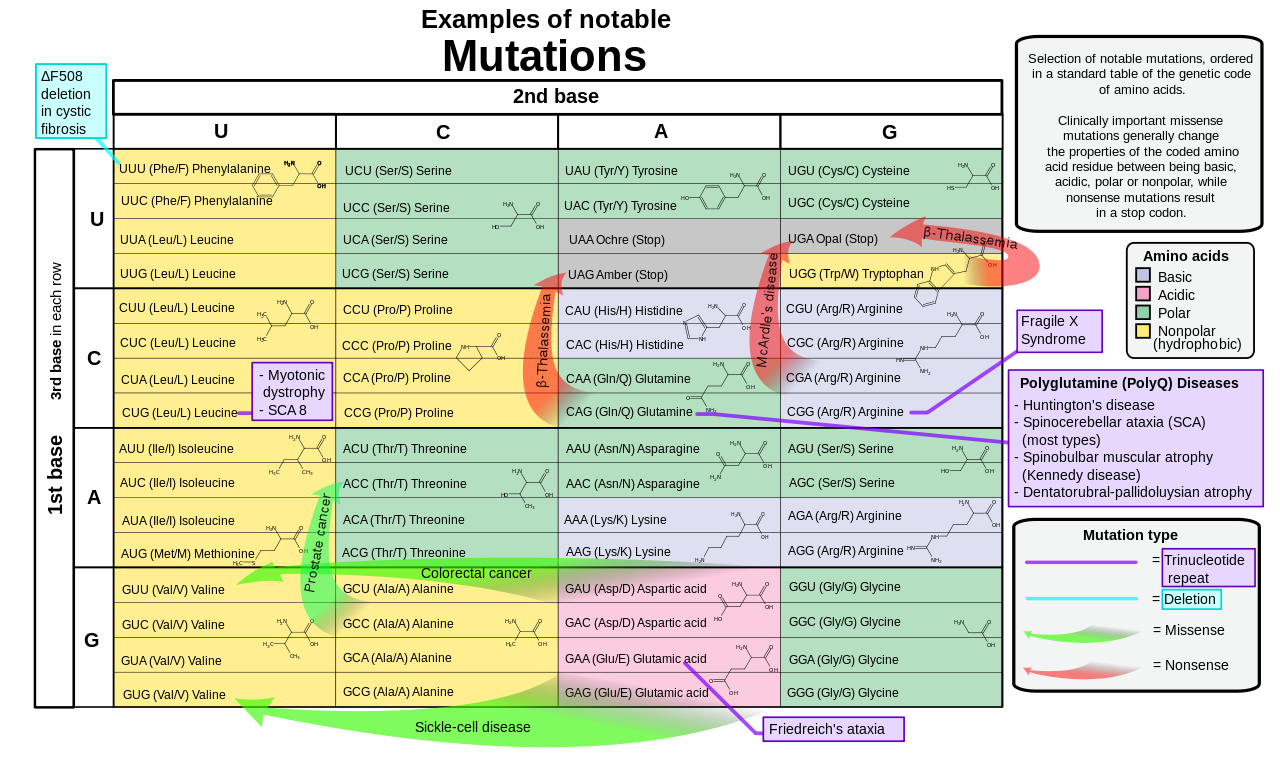
Selection of disease-causing mutations, in a standard table of the genetic code of amino acids.
Small-scale mutations affect a gene in one or a few nucleotides. (If only a single nucleotide is affected, they are called point mutations.) Small-scale mutations include:
Insertions add one or more extra nucleotides into the DNA. They are usually caused by transposable elements, or errors during replication of repeating elements. Insertions in the coding region of a gene may alter splicing of the mRNA (splice site mutation), or cause a shift in the reading frame (frameshift), both of which can significantly alter the gene product. Insertions can be reversed by excision of the transposable element.
Deletions remove one or more nucleotides from the DNA. Like insertions, these mutations can alter the reading frame of the gene. In general, they are irreversible: Though exactly the same sequence might, in theory, be restored by an insertion, transposable elements able to revert a very short deletion (say 1–2 bases) in any location either are highly unlikely to exist or do not exist at all.
Substitution mutations, often caused by chemicals or malfunction of DNA replication, exchange a single nucleotide for another. These changes are classified as transitions or transversions. Most common is the transition that exchanges a purine for a purine (A ↔ G) or a pyrimidine for a pyrimidine, (C ↔ T). A transition can be caused by nitrous acid, base mispairing, or mutagenic base analogs such as BrdU. Less common is a transversion, which exchanges a purine for a pyrimidine or a pyrimidine for a purine (C/T ↔ A/G). An example of a transversion is the conversion of adenine (A) into a cytosine (C). Point mutations are modifications of single base pairs of DNA or other small base pairs within a gene. A point mutation can be reversed by another point mutation, in which the nucleotide is changed back to its original state (true reversion) or by second-site reversion (a complementary mutation elsewhere that results in regained gene functionality). As discussed below, point mutations that occur within the protein coding region of a gene may be classified as synonymous or nonsynonymous substitutions, the latter of which in turn can be divided into missense or nonsense mutations.
The effect of a mutation on protein sequence depends in part on where in the genome it occurs, especially whether it is in a coding or non-coding region. Mutations in the non-coding regulatory sequences of a gene, such as promoters, enhancers, and silencers, can alter levels of gene expression, but are less likely to alter the protein sequence. Mutations within introns and in regions with no known biological function (e.g. pseudogenes,retrotransposons) are generally neutral, having no effect on phenotype – though intron mutations could alter the protein product if they affect mRNA splicing.
Mutations in that occur in coding regions of the genome are more likely to alter the protein product, and can be categorized by their effect on amino acid sequence:
A frameshift mutation is caused by insertion or deletion of a number of nucleotides that is not evenly divisible by three from a DNA sequence. Due to the triplet nature of gene expression by codons, the insertion or deletion can disrupt the reading frame, or the grouping of the codons, resulting in a completely different translation from the original.The earlier in the sequence the deletion or insertion occurs, the more altered the protein produced is. (For example, the code CCU GAC UAC CUA codes for the amino acids proline, aspartic acid, tyrosine, and leucine. If the U in CCU was deleted, the resulting sequence would be CCG ACU ACC UAx, which would instead code for proline, threonine, threonine, and part of another amino acid or perhaps a stop codon (where the x stands for the following nucleotide).) By contrast, any insertion or deletion that is evenly divisible by three is termed an in-frame mutation.
A point substitution mutation results in a change in a single nucleotide and can be either synonymous or nonsynonymous.
A synonymous substitution replaces a codon with another codon that codes for the same amino acid, so that the produced amino acid sequence is not modified. Synonymous mutations occur due to the degenerate nature of the genetic code. If this mutation does not result in any phenotypic effects, then it is called silent, but not all synonymous substitutions are silent. (There can also be silent mutations in nucleotides outside of the coding regions, such as the introns, because the exact nucleotide sequence is not as crucial as it is in the coding regions, but these are not considered synonymous substitutions.)
A nonsynonymous substitution replaces a codon with another codon that codes for a different amino acid, so that the produced amino acid sequence is modified. Nonsynonymous substitutions can be classified as nonsense or missense mutations:
A missense mutation changes a nucleotide to cause substitution of a different amino acid. This in turn can render the resulting protein nonfunctional. Such mutations are responsible for diseases such as Epidermolysis bullosa,sickle-cell disease, and SOD1-mediated ALS. On the other hand, if a missense mutation occurs in an amino acid codon that results in the use of a different, but chemically similar, amino acid, then sometimes little or no change is rendered in the protein. For example, a change from AAA to AGA will encode arginine, a chemically similar molecule to the intended lysine. In this latter case the mutation will have little or no effect on phenotype and therefore be neutral.
A nonsense mutation is a point mutation in a sequence of DNA that results in a premature stop codon, or a nonsense codon in the transcribed mRNA, and possibly a truncated, and often nonfunctional protein product. This sort of mutation has been linked to different mutations, such as congenital adrenal hyperplasia. (See Stop codon.)
Loss-of-function mutations, also called inactivating mutations, result in the gene product having less or no function (being partially or wholly inactivated). When the allele has a complete loss of function (null allele), it is often called an amorph or amorphic mutation in the Muller's morphs schema. Phenotypes associated with such mutations are most often recessive. Exceptions are when the organism is haploid, or when the reduced dosage of a normal gene product is not enough for a normal phenotype (this is called haploinsufficiency).
Gain-of-function mutations, also called activating mutations, change the gene product such that its effect gets stronger (enhanced activation) or even is superseded by a different and abnormal function. When the new allele is created, a heterozygote containing the newly created allele as well as the original will express the new allele; genetically this defines the mutations as dominant phenotypes. Several of Muller's morphs correspond to gain of function, including hypermorph (increased gene expression) and neomorph (novel function). In December 2017, the U.S. government lifted a temporary ban implemented in 2014 that banned federal funding for any new "gain-of-function" experiments that enhance pathogens "such as Avian influenza, SARS and the Middle East Respiratory Syndrome or MERS viruses."
Dominant negative mutations (also called antimorphic mutations) have an altered gene product that acts antagonistically to the wild-type allele. These mutations usually result in an altered molecular function (often inactive) and are characterized by a dominant or semi-dominant phenotype. In humans, dominant negative mutations have been implicated in cancer (e.g., mutations in genes p53,ATM,CEBPA and PPARgamma).Marfan syndrome is caused by mutations in the FBN1 gene, located on chromosome 15, which encodes fibrillin-1, a glycoprotein component of the extracellular matrix. Marfan syndrome is also an example of dominant negative mutation and haploinsufficiency.
Hypomorphs, after Mullerian classification, are characterized by altered gene products that acts with decreased gene expression compared to the wild type allele. Usually, hypomorphic mutations are recessive, but haploinsufficiency causes some alleles to be dominant.
Neomorphs are characterized by the control of new protein product synthesis.
Lethal mutations are mutations that lead to the death of the organisms that carry the mutations.
A back mutation or reversion is a point mutation that restores the original sequence and hence the original phenotype.
In applied genetics, it is usual to speak of mutations as either harmful or beneficial.
A harmful, or deleterious, mutation decreases the fitness of the organism.
A beneficial, or advantageous mutation increases the fitness of the organism.
A neutral mutation has no harmful or beneficial effect on the organism. Such mutations occur at a steady rate, forming the basis for the molecular clock. In the neutral theory of molecular evolution, neutral mutations provide genetic drift as the basis for most variation at the molecular level.
A nearly neutral mutation is a mutation that may be slightly deleterious or advantageous, although most nearly neutral mutations are slightly deleterious.
Attempts have been made to infer the distribution of fitness effects (DFE) using mutagenesis experiments and theoretical models applied to molecular sequence data. DFE, as used to determine the relative abundance of different types of mutations (i.e., strongly deleterious, nearly neutral or advantageous), is relevant to many evolutionary questions, such as the maintenance of genetic variation, the rate of genomic decay, the maintenance of outcrossingsexual reproduction as opposed to inbreeding and the evolution of sex and genetic recombination. DFE can also be tracked by tracking the skewness of the distribution of mutations with putatively severe effects as compared to the distribution of mutations with putatively mild or absent effect. In summary, the DFE plays an important role in predicting evolutionary dynamics. A variety of approaches have been used to study the DFE, including theoretical, experimental and analytical methods.
Mutagenesis experiment: The direct method to investigate the DFE is to induce mutations and then measure the mutational fitness effects, which has already been done in viruses, bacteria, yeast, and Drosophila. For example, most studies of the DFE in viruses used site-directed mutagenesis to create point mutations and measure relative fitness of each mutant. In Escherichia coli, one study used transposon mutagenesis to directly measure the fitness of a random insertion of a derivative of Tn10. In yeast, a combined mutagenesis and deep sequencing approach has been developed to generate high-quality systematic mutant libraries and measure fitness in high throughput. However, given that many mutations have effects too small to be detected and that mutagenesis experiments can detect only mutations of moderately large effect; DNA sequence data analysis can provide valuable information about these mutations.
Molecular sequence analysis: With rapid development of DNA sequencing technology, an enormous amount of DNA sequence data is available and even more is forthcoming in the future. Various methods have been developed to infer the DFE from DNA sequence data. By examining DNA sequence differences within and between species, we are able to infer various characteristics of the DFE for neutral, deleterious and advantageous mutations. To be specific, the DNA sequence analysis approach allows us to estimate the effects of mutations with very small effects, which are hardly detectable through mutagenesis experiments.
One of the earliest theoretical studies of the distribution of fitness effects was done by Motoo Kimura, an influential theoretical population geneticist. His neutral theory of molecular evolution proposes that most novel mutations will be highly deleterious, with a small fraction being neutral. Hiroshi Akashi more recently proposed a bimodal model for the DFE, with modes centered around highly deleterious and neutral mutations. Both theories agree that the vast majority of novel mutations are neutral or deleterious and that advantageous mutations are rare, which has been supported by experimental results. One example is a study done on the DFE of random mutations in vesicular stomatitis virus. Out of all mutations, 39.6% were lethal, 31.2% were non-lethal deleterious, and 27.1% were neutral. Another example comes from a high throughput mutagenesis experiment with yeast. In this experiment it was shown that the overall DFE is bimodal, with a cluster of neutral mutations, and a broad distribution of deleterious mutations.
Though relatively few mutations are advantageous, those that are play an important role in evolutionary changes. Like neutral mutations, weakly selected advantageous mutations can be lost due to random genetic drift, but strongly selected advantageous mutations are more likely to be fixed. Knowing the DFE of advantageous mutations may lead to increased ability to predict the evolutionary dynamics. Theoretical work on the DFE for advantageous mutations has been done by John H. Gillespie and H. Allen Orr. They proposed that the distribution for advantageous mutations should be exponential under a wide range of conditions, which, in general, has been supported by experimental studies, at least for strongly selected advantageous mutations.
In general, it is accepted that the majority of mutations are neutral or deleterious, with advantageous mutations being rare; however, the proportion of types of mutations varies between species. This indicates two important points: first, the proportion of effectively neutral mutations is likely to vary between species, resulting from dependence on effective population size; second, the average effect of deleterious mutations varies dramatically between species. In addition, the DFE also differs between coding regions and noncoding regions, with the DFE of noncoding DNA containing more weakly selected mutations.
A mutation has caused this moss rose plant to produce flowers of different colors. This is a somatic mutation that may also be passed on in the germline.
In multicellular organisms with dedicated reproductive cells, mutations can be subdivided into germline mutations, which can be passed on to descendants through their reproductive cells, and somatic mutations (also called acquired mutations), which involve cells outside the dedicated reproductive group and which are not usually transmitted to descendants.
Diploid organisms (e.g., humans) contain two copies of each gene—a paternal and a maternal allele. Based on the occurrence of mutation on each chromosome, we may classify mutations into three types. A wild type or homozygous non-mutated organism is one in which neither allele is mutated.
A heterozygous mutation is a mutation of only one allele.
A homozygous mutation is an identical mutation of both the paternal and maternal alleles.
Compound heterozygous mutations or a genetic compound consists of two different mutations in the paternal and maternal alleles.
A germline mutation in the reproductive cells of an individual gives rise to a constitutional mutation in the offspring, that is, a mutation that is present in every cell. A constitutional mutation can also occur very soon after fertilisation, or continue from a previous constitutional mutation in a parent. A germline mutation can be passed down through subsequent generations of organisms.
The distinction between germline and somatic mutations is important in animals that have a dedicated germline to produce reproductive cells. However, it is of little value in understanding the effects of mutations in plants, which lack a dedicated germline. The distinction is also blurred in those animals that reproduce asexually through mechanisms such as budding, because the cells that give rise to the daughter organisms also give rise to that organism's germline.
A new germline mutation not inherited from either parent is called a de novo mutation.
A change in the genetic structure that is not inherited from a parent, and also not passed to offspring, is called a somatic mutation. Somatic mutations are not inherited by an organism's offspring because they do not affect the germline. However, they are passed down to all the progeny of a mutated cell within the same organism during mitosis. A major section of an organism therefore might carry the same mutation. These types of mutations are usually prompted by environmental causes, such as ultraviolet radiation or any exposure to certain harmful chemicals, and can cause diseases including cancer.
With plants, some somatic mutations can be propagated without the need for seed production, for example, by grafting and stem cuttings. These type of mutation have led to new types of fruits, such as the "Delicious" apple and the "Washington" navel orange
Human and mouse somatic cells have a mutation rate more than ten times higher than the germline mutation rate for both species; mice have a higher rate of both somatic and germline mutations per cell division than humans. The disparity in mutation rate between the germline and somatic tissues likely reflects the greater importance of genome maintenance in the germline than in the soma.
Conditional mutation is a mutation that has wild-type (or less severe) phenotype under certain "permissive" environmental conditions and a mutant phenotype under certain "restrictive" conditions. For example, a temperature-sensitive mutation can cause cell death at high temperature (restrictive condition), but might have no deleterious consequences at a lower temperature (permissive condition). These mutations are non-autonomous, as their manifestation depends upon presence of certain conditions, as opposed to other mutations which appear autonomously. The permissive conditions may be temperature, certain chemicals, light or mutations in other parts of the genome.Invivo mechanisms like transcriptional switches can create conditional mutations. For instance, association of Steroid Binding Domain can create a transcriptional switch that can change the expression of a gene based on the presence of a steroid ligand. Conditional mutations have applications in research as they allow control over gene expression. This is especially useful studying diseases in adults by allowing expression after a certain period of growth, thus eliminating the deleterious effect of gene expression seen during stages of development in model organisms. DNA Recombinase systems like Cre-Lox recombination used in association with promoters that are activated under certain conditions can generate conditional mutations. Dual Recombinase technology can be used to induce multiple conditional mutations to study the diseases which manifest as a result of simultaneous mutations in multiple genes. Certain inteins have been identified which splice only at certain permissive temperatures, leading to improper protein synthesis and thus, loss-of-function mutations at other temperatures. Conditional mutations may also be used in genetic studies associated with ageing, as the expression can be changed after a certain time period in the organism's lifespan.
In order to categorize a mutation as such, the "normal" sequence must be obtained from the DNA of a "normal" or "healthy" organism (as opposed to a "mutant" or "sick" one), it should be identified and reported; ideally, it should be made publicly available for a straightforward nucleotide-by-nucleotide comparison, and agreed upon by the scientific community or by a group of expert geneticists and biologists, who have the responsibility of establishing the standard or so-called "consensus" sequence. This step requires a tremendous scientific effort. Once the consensus sequence is known, the mutations in a genome can be pinpointed, described, and classified. The committee of the Human Genome Variation Society (HGVS) has developed the standard human sequence variant nomenclature, which should be used by researchers and DNA diagnostic centers to generate unambiguous mutation descriptions. In principle, this nomenclature can also be used to describe mutations in other organisms. The nomenclature specifies the type of mutation and base or amino acid changes.
Nucleotide substitution (e.g., 76A>T) – The number is the position of the nucleotide from the 5' end; the first letter represents the wild-type nucleotide, and the second letter represents the nucleotide that replaced the wild type. In the given example, the adenine at the 76th position was replaced by a thymine.
If it becomes necessary to differentiate between mutations in genomic DNA,mitochondrial DNA, and RNA, a simple convention is used. For example, if the 100th base of a nucleotide sequence mutated from G to C, then it would be written as g.100G>C if the mutation occurred in genomic DNA, m.100G>C if the mutation occurred in mitochondrial DNA, or r.100g>c if the mutation occurred in RNA. Note that, for mutations in RNA, the nucleotide code is written in lower case.
Amino acid substitution (e.g., D111E) – The first letter is the one letter code of the wild-type amino acid, the number is the position of the amino acid from the N-terminus, and the second letter is the one letter code of the amino acid present in the mutation. Nonsense mutations are represented with an X for the second amino acid (e.g. D111X).
Amino acid deletion (e.g., ΔF508) – The Greek letter Δ (delta) indicates a deletion. The letter refers to the amino acid present in the wild type and the number is the position from the N terminus of the amino acid were it to be present as in the wild type.
Mutation rates vary substantially across species, and the evolutionary forces that generally determine mutation are the subject of ongoing investigation.
In humans, the mutation rate is about 50-90 de novo mutations per genome per generation, that is, each human accumulates about 50-90 novel mutations that were not present in his or her parents. This number has been established by sequencing thousands of human trios, that is, two parents and at least one child.
The genomes of RNA viruses are based on RNA rather than DNA. The RNA viral genome can be double-stranded (as in DNA) or single-stranded. In some of these viruses (such as the single-stranded human immunodeficiency virus), replication occurs quickly, and there are no mechanisms to check the genome for accuracy. This error-prone process often results in mutations.
Disease causation
Disease causation
Disease causation (W)
Changes in DNA caused by mutation in a coding region of DNA can cause errors in protein sequence that may result in partially or completely non-functional proteins. Each cell, in order to function correctly, depends on thousands of proteins to function in the right places at the right times. When a mutation alters a protein that plays a critical role in the body, a medical condition can result. One study on the comparison of genes between different species of Drosophila suggests that if a mutation does change a protein, the mutation will most likely be harmful, with an estimated 70 percent of amino acid polymorphisms having damaging effects, and the remainder being either neutral or weakly beneficial. Some mutations alter a gene's DNA base sequence but do not change the protein made by the gene. Studies have shown that only 7% of point mutations in noncoding DNA of yeast are deleterious and 12% in coding DNA are deleterious. The rest of the mutations are either neutral or slightly beneficial.
If a mutation is present in a germ cell, it can give rise to offspring that carries the mutation in all of its cells. This is the case in hereditary diseases. In particular, if there is a mutation in a DNA repair gene within a germ cell, humans carrying such germline mutations may have an increased risk of cancer. A list of 34 such germline mutations is given in the article DNA repair-deficiency disorder. An example of one is albinism, a mutation that occurs in the OCA1 or OCA2 gene. Individuals with this disorder are more prone to many types of cancers, other disorders and have impaired vision.
DNA damage can cause an error when the DNA is replicated, and this error of replication can cause a gene mutation that, in turn, could cause a genetic disorder. DNA damages are repaired by the DNA repair system of the cell. Each cell has a number of pathways through which enzymes recognize and repair damages in DNA. Because DNA can be damaged in many ways, the process of DNA repair is an important way in which the body protects itself from disease. Once DNA damage has given rise to a mutation, the mutation cannot be repaired.
On the other hand, a mutation may occur in a somatic cell of an organism. Such mutations will be present in all descendants of this cell within the same organism. The accumulation of certain mutations over generations of somatic cells is part of cause of malignant transformation, from normal cell to cancer cell.
Cells with heterozygous loss-of-function mutations (one good copy of gene and one mutated copy) may function normally with the unmutated copy until the good copy has been spontaneously somatically mutated. This kind of mutation happens often in living organisms, but it is difficult to measure the rate. Measuring this rate is important in predicting the rate at which people may develop cancer.
Point mutations may arise from spontaneous mutations that occur during DNA replication. The rate of mutation may be increased by mutagens. Mutagens can be physical, such as radiation from UV rays,X-rays or extreme heat, or chemical (molecules that misplace base pairs or disrupt the helical shape of DNA). Mutagens associated with cancers are often studied to learn about cancer and its prevention.
Prion mutations
Prion mutations (W) Prions are proteins and do not contain genetic material. However, prion replication has been shown to be subject to mutation and natural selection just like other forms of replication. The human gene PRNP codes for the major prion protein, PrP, and is subject to mutations that can give rise to disease-causing prions.
Although mutations that cause changes in protein sequences can be harmful to an organism, on occasions the effect may be positive in a given environment. In this case, the mutation may enable the mutant organism to withstand particular environmental stresses better than wild-type organisms, or reproduce more quickly. In these cases a mutation will tend to become more common in a population through natural selection. Examples include the following:
HIV resistance: a specific 32 base pair deletion in human CCR5 (CCR5-Δ32) confers HIV resistance to homozygotes and delays AIDS onset in heterozygotes. One possible explanation of the etiology of the relatively high frequency of CCR5-Δ32 in the European population is that it conferred resistance to the bubonic plague in mid-14th century Europe. People with this mutation were more likely to survive infection; thus its frequency in the population increased. This theory could explain why this mutation is not found in Southern Africa, which remained untouched by bubonic plague. A newer theory suggests that the selective pressure on the CCR5 Delta 32 mutation was caused by smallpox instead of the bubonic plague.
Malaria resistance: An example of a harmful mutation is sickle-cell disease, a blood disorder in which the body produces an abnormal type of the oxygen-carrying substance hemoglobin in the red blood cells. One-third of all indigenous inhabitants of Sub-Saharan Africa carry the allele, because, in areas where malaria is common, there is a survival value in carrying only a single sickle-cell allele (sickle cell trait). Those with only one of the two alleles of the sickle-cell disease are more resistant to malaria, since the infestation of the malaria Plasmodium is halted by the sickling of the cells that it infests.
Antibiotic resistance: Practically all bacteria develop antibiotic resistance when exposed to antibiotics. In fact, bacterial populations already have such mutations that get selected under antibiotic selection. Obviously, such mutations are only beneficial for the bacteria but not for those infected.
Lactase persistence. A mutation allowed humans to express the enzyme lactase after they are naturally weaned from breast milk, allowing adults to digest lactose, which is likely one of the most beneficial mutations in recent human evolution.
Mutationism is one of several alternatives to evolution by natural selection that have existed both before and after the publication of Charles Darwin's 1859 book, On the Origin of Species. In the theory, mutation was the source of novelty, creating new forms and new species, potentially instantaneously, in a sudden jump. This was envisaged as driving evolution, which was limited by the supply of mutations.
Before Darwin, biologists commonly believed in saltationism, the possibility of large evolutionary jumps, including immediate speciation. For example, in 1822 Étienne Geoffroy Saint-Hilaire argued that species could be formed by sudden transformations, or what would later be called macromutation. Darwin opposed saltation, insisting on gradualism in evolution as in geology. In 1864, Albert von Kölliker revived Geoffroy's theory. In 1901 the geneticistHugo de Vries gave the name "mutation" to seemingly new forms that suddenly arose in his experiments on the evening primrose Oenothera lamarckiana, and in the first decade of the 20th century, mutationism, or as de Vries named it mutationstheorie, became a rival to Darwinism supported for a while by geneticists including William Bateson,Thomas Hunt Morgan, and Reginald Punnett.
Understanding of mutationism is clouded by the mid-20th century portrayal of the early mutationists by supporters of the modern synthesis as opponents of Darwinian evolution and rivals of the biometrics school who argued that selection operated on continuous variation. In this portrayal, mutationism was defeated by a synthesis of genetics and natural selection that supposedly started later, around 1918, with work by the mathematician Ronald Fisher. However, the alignment of Mendelian genetics and natural selection began as early as 1902 with a paper by Udny Yule, and built up with theoretical and experimental work in Europe and America. Despite the controversy, the early mutationists had by 1918 already accepted natural selection and explained continuous variation as the result of multiple genes acting on the same characteristic, such as height.
Mutationism, along with other alternatives to Darwinism like Lamarckism and orthogenesis, was discarded by most biologists as they came to see that Mendelian genetics and natural selection could readily work together; mutation took its place as a source of the genetic variation essential for natural selection to work on. However, mutationism did not entirely vanish. In 1940, Richard Goldschmidt again argued for single-step speciation by macromutation, describing the organisms thus produced as "hopeful monsters", earning widespread ridicule. In 1987, Masatoshi Nei argued controversially that evolution was often mutation-limited. Modern biologists such as Douglas J. Futuyma conclude that essentially all claims of evolution driven by large mutations can be explained by Darwinian evolution.


.png)


.jpg)