Örgenliğin genetik planı DNAnın nükleotidlerinde bulunur.
Örgenliğin genetik planı DNAnın eşyazımları olan RNA molekülleri tarafından yerine getirilir.
RNA genetik bilgileri DNAdan ribozomlara taşımaktan daha çoğunu yapar.
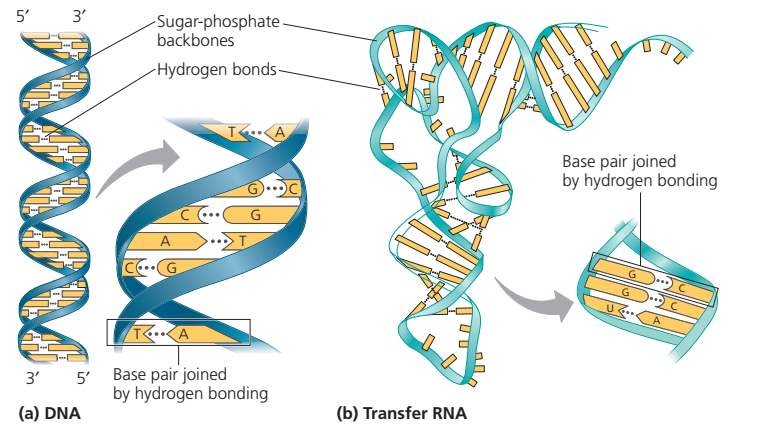
Eşyazım (transcription) DNAnın tek teline tümleyici RNAnın üretimidir (A T ile, U A ile, G C ile, ve C G ile).
Eşyazım sürecini RNA polimeraz yönetir.
🛑 RNA YAPISI
RNA YAPISI
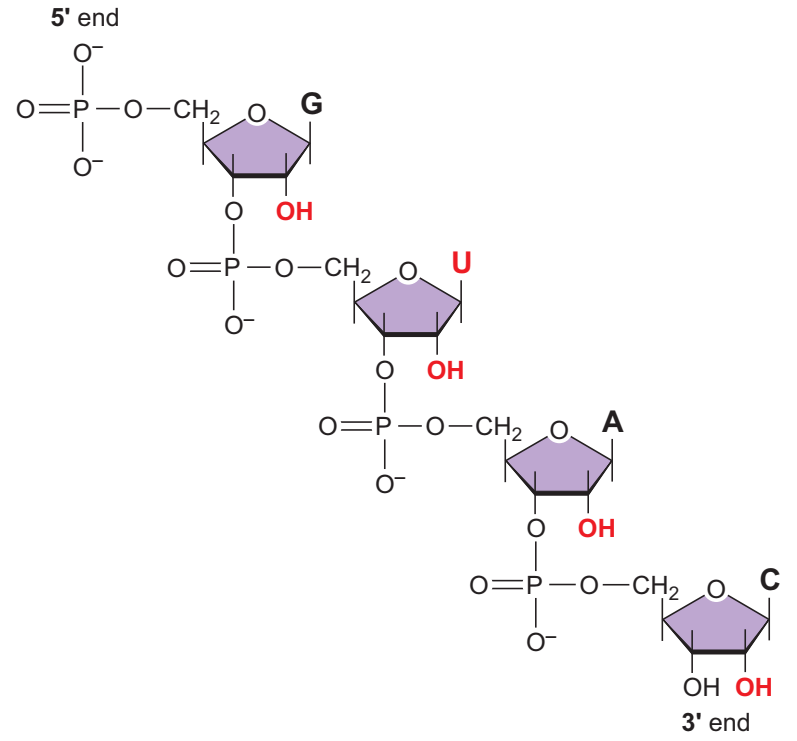
Kısa bir RNA molekülü. Nükleotidler arasındaki fosfodiester bağ DNAdaki ile aynıdır.
RNA da DNA gibi bir nükleotid zinciridir ve boyu birkaç bin nükleotid kadar ya da çok daha kısadır.
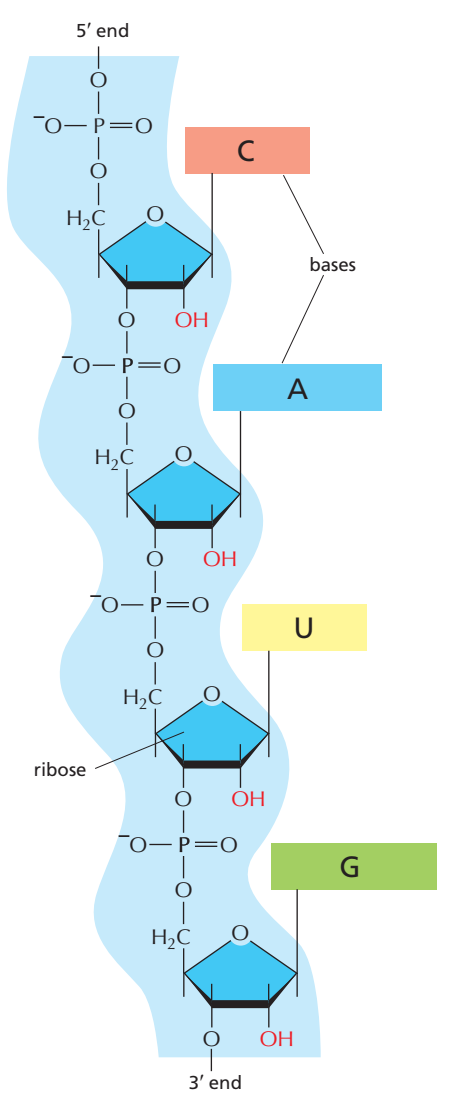
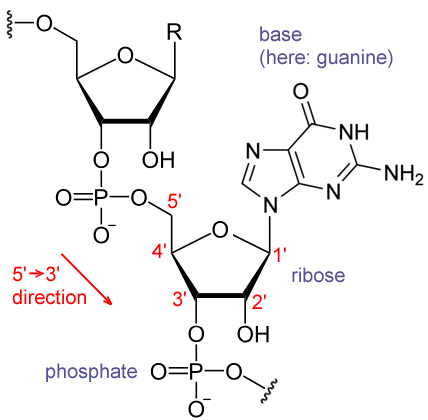
RNAda nükleotidler fosfodiester bağı ile birbirine eklenerek 5ʹ—3ʹ yönünde doğrusal bir zincir oluşturur.
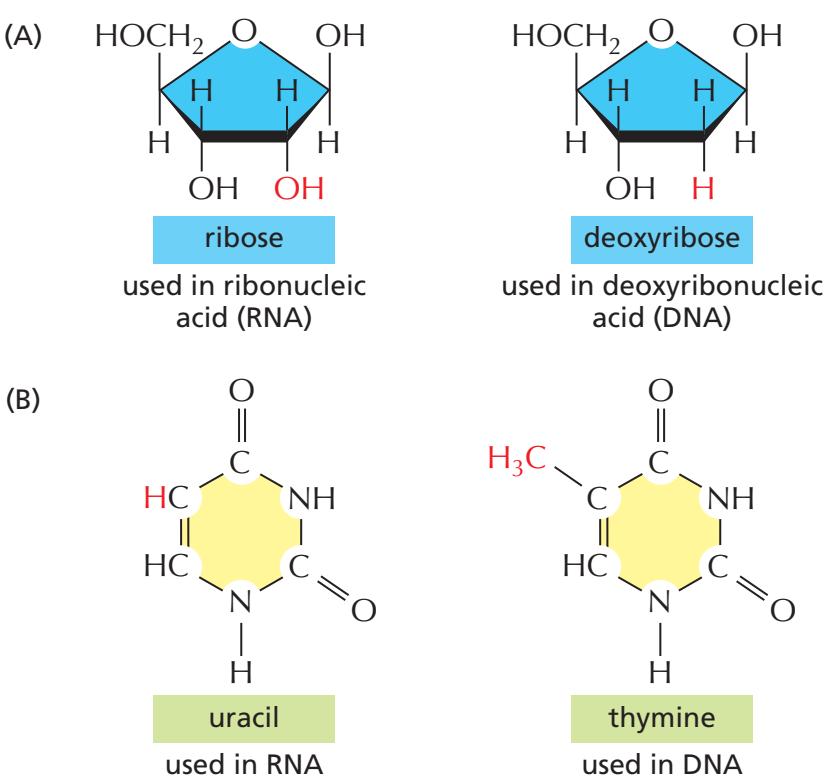
(A) RNA DNAda kullanılan deoksiribozdan bir ek —OH grubu ile ayrılan riboz şekerini kullanır.
(B) RNA DNAda kullanılan thimin bazı yerine ondan bir —CH3 grubunun yokluğu ile ayrılan urasil bazını kullanır.
RNAnın çift sarmal karakteristikleri.
Tümleyici dizileri olan bir RNA molekülünde,
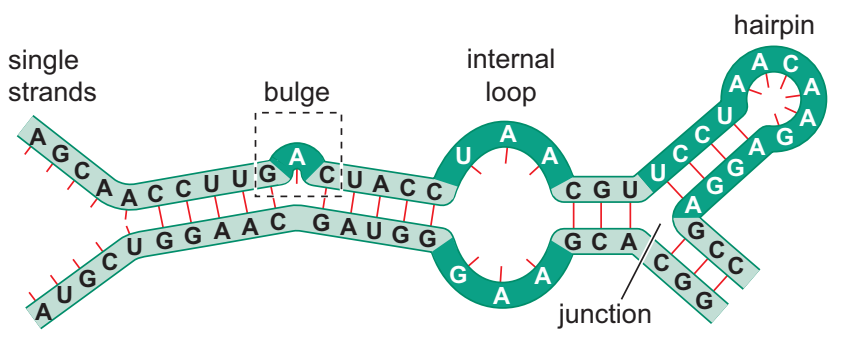
araya giren RNA kesimleri (tümleyici-olmayan) şekilde örneklenen yapıları oluşturmak üzere dışarıya doğru ilmeklenebilir: Bir tümsek (bulge), bir iç ilmek (internal loop), ya da bir toka (hairpin).
Çift telli nükleik asidin boyu baz çiftlerinin sayısı ile ölçülür.
Tek telli nükleik asidin boyu nükleotidlerin sayısı ile ölçülür (kilobaz: Kb; megabaz: Mb).
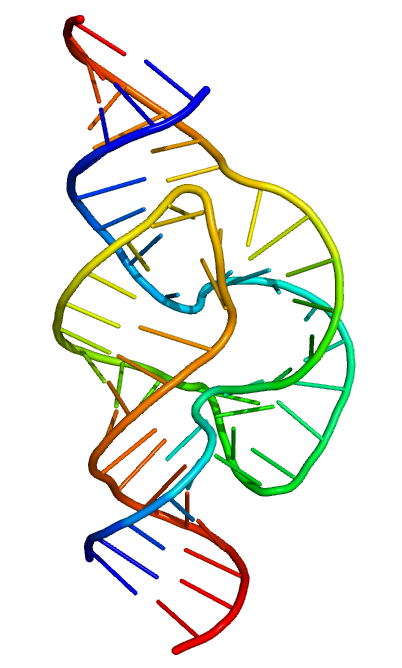
RNA DNA gibi sarmal yapı biçiminde değil, kendi üzerine katlanan tekil zincir olarak bulunur.
RNAnın çift sarmal karakteristikleri.
Tümleyici dizileri olan bir RNA molekülünde,
araya giren RNA kesimleri (tümleyici-olmayan) şekilde örneklenen yapıları oluşturmak üzere dışarıya doğru ilmeklenebilir: Bir tümsek (bulge), bir iç ilmek (internal loop), ya da bir toka (hairpin).
RNA genlerin kodlama, kod çözme, düzenleme ve anlatımında özsel işlevler yerine getirir.
RNA azot bazları olarak Guanin, Urasil, Adenin ve Sitozin kullanır (G, U, A ve C).
RNAyı DNAdan ayırdeden etmen omurgada şeker bölümü olarak deoksiriboz yerine riboz, ve nükleotid thimin (T) yerine urasil (U) kullanmasıdır (A, C, G, T ve U ölçün nükleotidlerdir).
RNAda G:C ve A:U tümleyici baz çiftlerdir ve G:U çifti de kararlıdır.
RNA dördüncül yapısı nedeniyle proteinler gibi kimyasal kataliz yapabilir (enzimler gibi).
Ribozimler olarak bilinen RNA enzimlerinde bir etkin site, bir özdek için bir bağlama sitesi, ve örneğin bir metal ion gibi bir kofaktör için bir bağlanma sitesi vardır.
Kimi işlevsel RNA molekülleri, özellikle tRNAlar ölçün olmayan nükleotidler kapsar (bunlar ölçün nükleotidlerin kimyasal değişkileri yoluyla RNA eşyazımından sonra oluşur).
RNA ÜRETİMİ (EŞYAZIM/TRANSCRIPTION)
RNA polimeraz enzimi DNAdan eşyazım yapar. RNA polimeraz (açık mavi) DNA boyunca adım adım ilerlerken kataliz için gerekli Mg2+ ile belirtilen etkin sitesinde (kırmızı) DNA sarmalını açar. İlerlerken, polimeraz açığa çıkan bir DNA telini kalıp olarak kullanarak polimerizasyon sitesinde nükleotidleri birer birer RNA zincirine ekler. RNA eşyazımı böylece iki DNA telinden birinin tümleyici eşlemidir. DNA/RNA sarmalının kısa bir bölgesi (yaklaşık dokuz nükleotid çifti uzunlukta) yalnızca geçici olarak oluşur, ve DNA/RNA sarmalının bir “penceresi” buna göre polimeraz ile DNA boyunca ilerler ve arkasında DNA çifte sarmalı yeniden kurulur. İçeri giren nükleotidler ribonükleosid trifosfat biçimindedir (ATP, UTP, CTP ve GTP), ve fosfat-fosfat bağlarında depolanan enerji polimerizasyon tepkimesi için itici gücü sağlar. Bir X-ışını kristalografik yapı üzerine dayanan figür polimerazın kesit bir görünüşünü temsil eder; iç bölümü göstermek için molekülün ön parçası çıkarıldı. (Adapted from P. Cramer et al., Science
288:640–649, 2000; PDB code: 1HQM.) (Molecular Biology of the Cell, 2017, s. 304)
RNA polimeraz molekülleri aynı DNA kalıbı üzerinde birbirini saniyede 50 nükleotid gibi bir hızla ilerleyerek RNA moleküllerini üretir.
RNA polimeraz RNAya eşlemlenen her 104 nükleotid için yaklaşık bir hata yapar ve yine kendi düzeltme düzeneği ile geri dönerek hatayı düzeltebilir (DNA durumunda oran 107 nükleotidde birdir).
Protein bireşimine katılan başlıca üç RNA tipi vardır:
İletmen RNA (mRNA);
Ribozomal RNA (rRNA);
Aktarıcı RNA (tRNA).
Bu üç RNA tipi, DNA gibi, dallanmayan polimerik moleküllerden oluşur.
Polimeri oluşturan nükleosid monofosfat monomerleri fosfodiester bağları tarafından biraraya tutturulur.
(Primerler oligonükleotidlerdir ve DNA kalıbına tümleyici olarak bağlanırlar ve DNA polimerazın uzatma yapmasını sağlarlar.
Primaz RNA primer bireşimi yapan enzimdir.
Primazı eşlenimin başına yükleyen özel proteinler böylelikle öncü DNA telinin bireşimi başlatırlar.
Sonraki adım daha başka eşlenim proteinlerinin primazı geciken karşı tel üzerinde DNA eşlenimini başlatmaya götürmesidir.
Hem öncü hem de geciken tel primerleri DNA polimerazlar tarafından uzatıldıktan sonra, RNA primerler enzimatik olarak çıkarılır ve DNA dizisinde doğan boşluk DNA polimeraz I ve DNA ligaz tarafından doldurulur.)
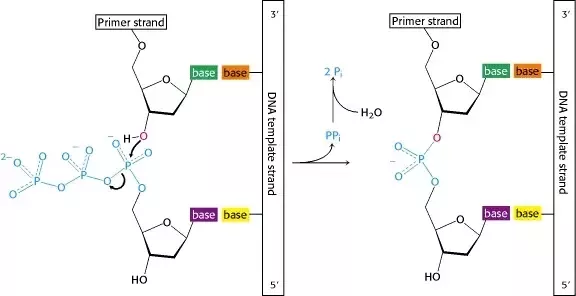
Nükleosidin 3'-hidroksili nükleosid trifosfatın 5' α-fosfatına saldırısı. Uzaklaştırılan öteki iki fosfat bir nükleotidde sonuçlanır.
DNA eşlenimi sırasında, DNA polimeraz III kullanılan birincil polimerazdır. Pol III yeni bireştirilen telin büyümesini katalize eder ve bunu bir nükleosidin (fosfatlar olmaksızın şeker + baz) 3'-hidroksilinin bitişik nükleosid trifosfatın 5'-α-fosfatına saldırısı ile yapar (α-fosfat şekerin en yakınında olandır). Sonuç bir nükleotidin polinükleotid zincire eklenmesidir.
📹📹📹 RNA (VİDEO)
📹 RNA and Transcription (updated) / Beverly Biology (VİDEO)
📹 RNA and Transcription (updated) / Beverly Biology (LINK)
Paul Andersen continues his description of DNA and RNA. He begins with the structure of DNA and RNA and moves into the process of DNA Replication. He also describes the central dogma of biology explaining how DNA is transcribed to mRNA and is finally translated into proteins. He also introduces genetic engineering and explains how transformation is used to create insulin.
📹 mRNA, tRNA, and rRNA function / Types of RNA 2min (VİDEO)
📹 mRNA, tRNA, and rRNA function / Types of RNA 2min (LINK)
DNA is a type of nucleic acid. And in this video we are going to have a quick look at another type; RNA.
We have both DNA and RNA in our bodies. We need them both! DNA is the blueprint; it contains all of the instructions for the cell to grow, function and replicate. The RNA carries out these instructions; it copies and transfers the genetic code from the DNA to ensure the relevant proteins are made. So just think of it as “DNA makes RNA make proteins”.
Whereas DNA stands for deoxyribonucleic acid, RNA stands for ribonucleic acid.
Whilst DNA is double stranded, RNA is single stranded.
Like DNA, RNA is made up of a long chain of nucleotides. Each nucleotide consists of a ribose sugar, phosphate group and a nucleotide base. RNA has a sugar called ribose, whereas DNA has a sugar called deoxyribose.
RNA has a base uracil, or U, whereas DNA has the base thymine or T. So in RNA C and G still pair, but now A and U pair.
We need to know about RNA because we will see it when learning about protein synthesis. We will see two special types of RNA: messenger RNA which is known as mRNA. And transfer RNA which is known as tRNA. We will see the mRNA being synthesised inside the nucleus, copied from the DNA code. The tRNA is found in the cytoplasm. For many years, we just thought RNA was a DNA photocopier as mRNA, the protein builder as tRNA and found in ribosomes as rRNA. However, RNA can also act as enzymes to speed up chemical reactions. And in many viruses, they have RNA instead of DNA. The RNA carries the genetic code in these viruses.
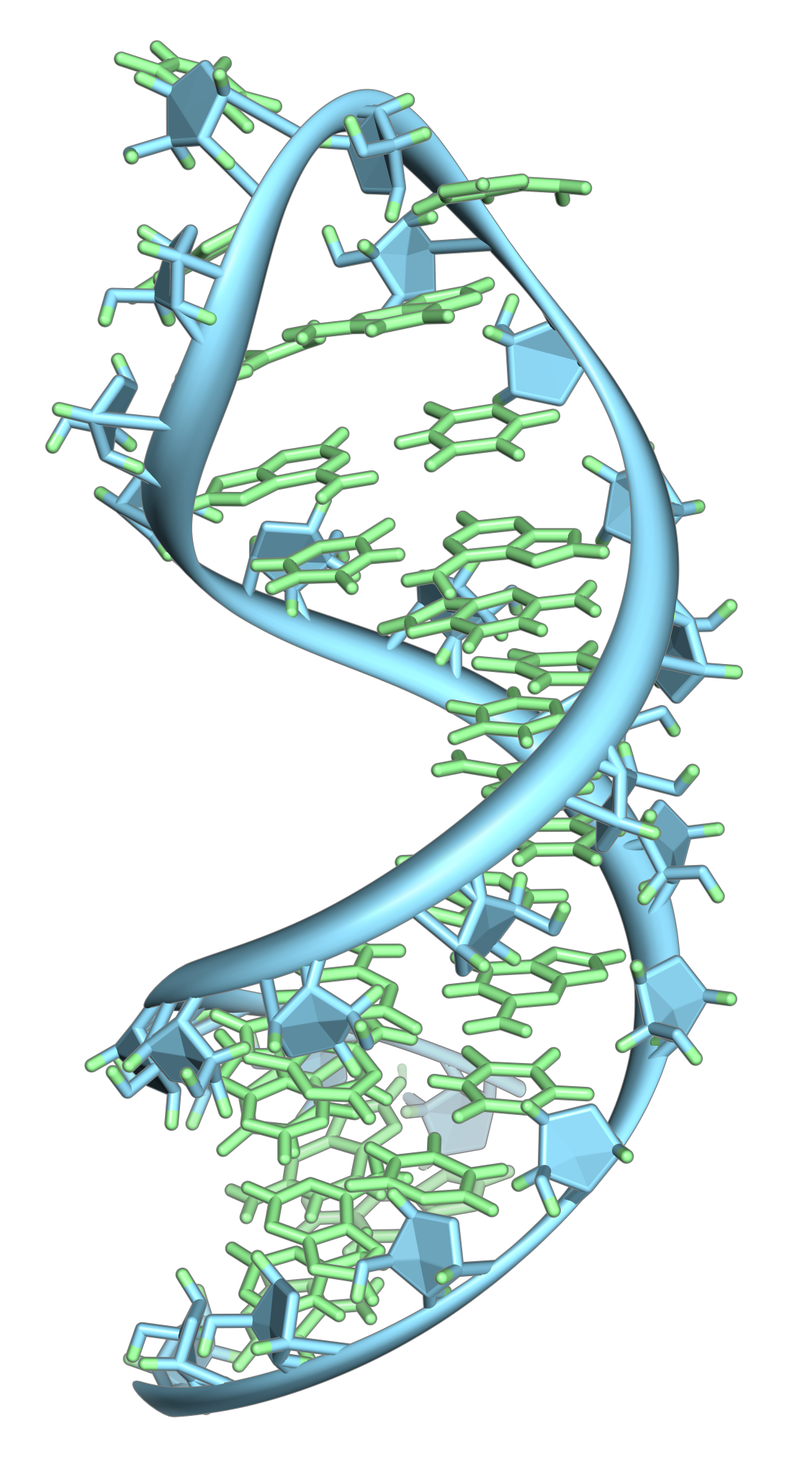
A hairpin loop from a pre-mRNA. Highlighted are the nucleobases (green) and the ribose-phosphate backbone (blue). This is a single strand of RNA that folds back upon itself.
Some RNA molecules play an active role within cells by catalyzing biological reactions, controlling gene expression, or sensing and communicating responses to cellular signals. One of these active processes is protein synthesis, a universal function in which RNA molecules direct the synthesis of proteins on ribosomes. This process uses transfer RNA (tRNA) molecules to deliver amino acids to the ribosome, where ribosomal RNA (rRNA) then links amino acids together to form coded proteins.
The chemical structure of RNA is very similar to that of DNA, but differs in three primary ways:
Unlike double-stranded DNA, RNA is a single-stranded molecule in many of its biological roles and consists of much shorter chains of nucleotides. However, a single RNA molecule can, by complementary base pairing, form intrastrand double helixes, as in tRNA.
While the sugar-phosphate "backbone" of DNA contains deoxyribose, RNA contains ribose instead. Ribose has a hydroxyl group attached to the pentose ring in the 2' position, whereas deoxyribose does not. The hydroxyl groups in the ribose backbone make RNA more chemically labile than DNA by lowering the activation energy of hydrolysis.
Like DNA, most biologically active RNAs, including mRNA,tRNA,rRNA,snRNAs, and other non-coding RNAs, contain self-complementary sequences that allow parts of the RNA to fold and pair with itself to form double helices. Analysis of these RNAs has revealed that they are highly structured. Unlike DNA, their structures do not consist of long double helices, but rather collections of short helices packed together into structures akin to proteins.
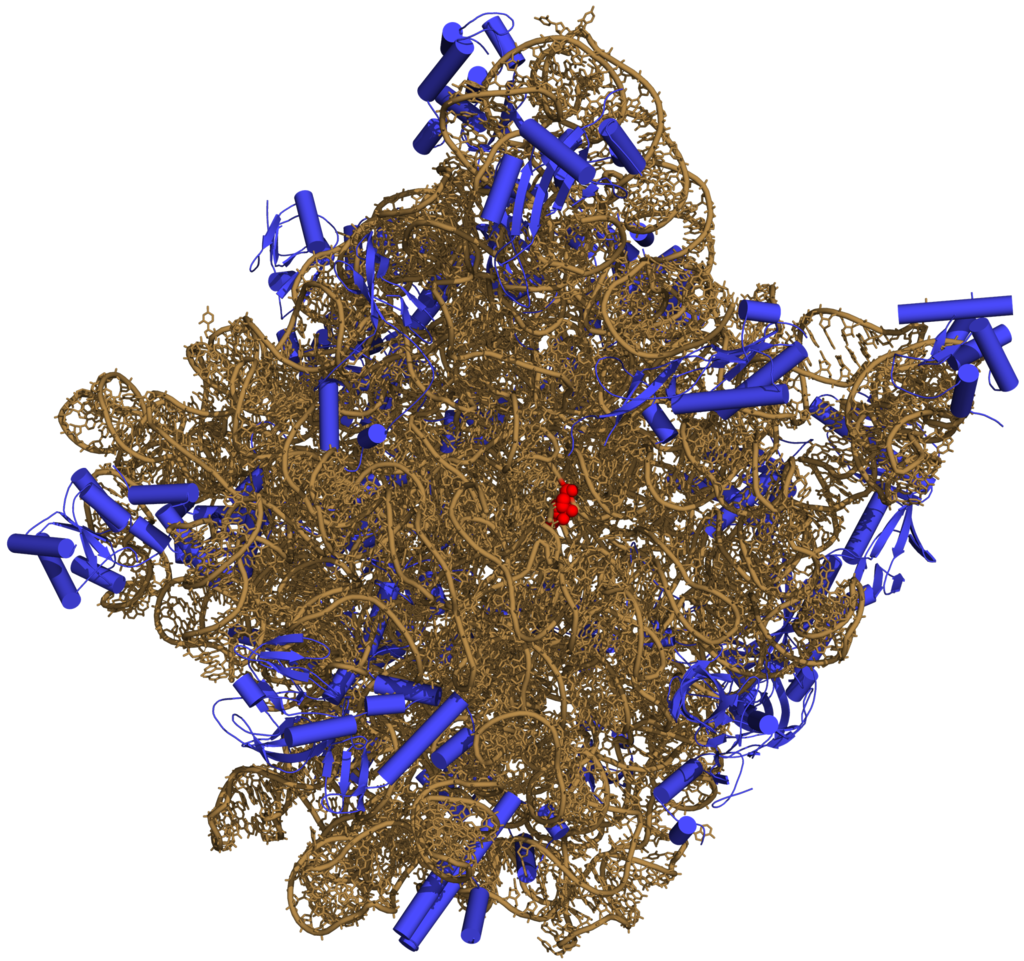
In this fashion, RNAs can achieve chemical catalysis (like enzymes). For instance, determination of the structure of the ribosome — an RNA-protein complex that catalyzes peptide bond formation — revealed that its active site is composed entirely of RNA.
Three-dimensional representation of the 50S ribosomal subunit. Ribosomal RNA is in ochre, proteins in blue. The active site is a small segment of rRNA, indicated in red.
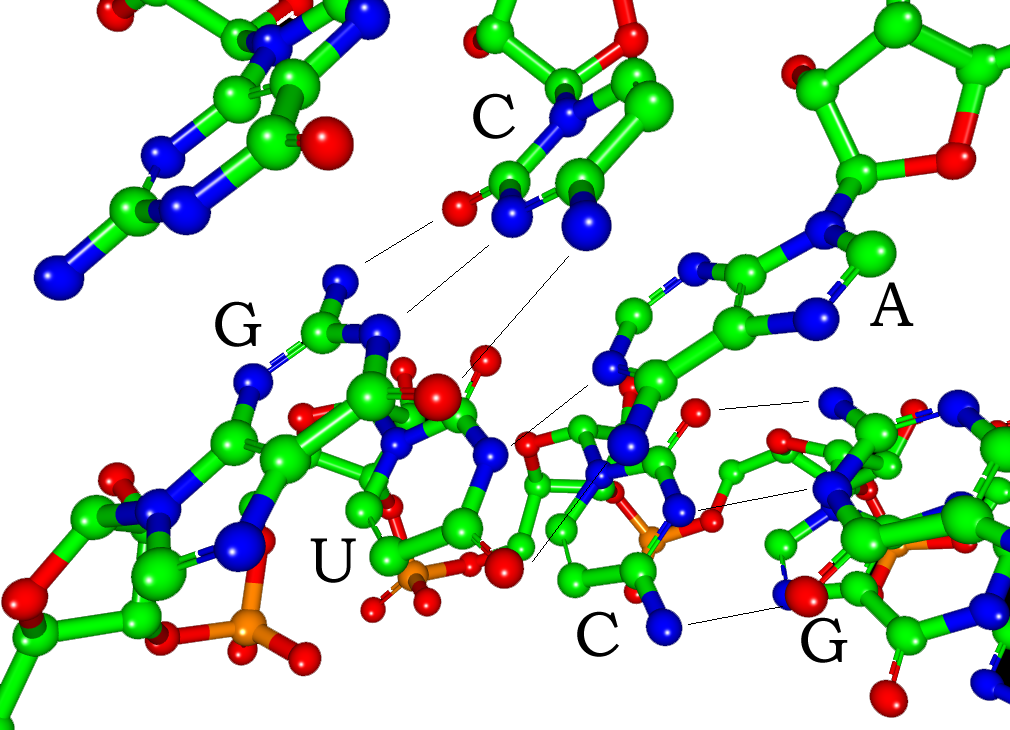
Each nucleotide in RNA contains a ribose sugar, with carbons numbered 1' through 5'. A base is attached to the 1' position, in general, adenine (A), cytosine (C), guanine (G), or uracil (U). Adenine and guanine are purines, cytosine and uracil are pyrimidines. A phosphate group is attached to the 3' position of one ribose and the 5' position of the next. The phosphate groups have a negative charge each, making RNA a charged molecule (polyanion).The bases form hydrogen bonds between cytosine and guanine, between adenine and uracil and between guanine and uracil. However, other interactions are possible, such as a group of adenine bases binding to each other in a bulge, or the GNRA tetraloop that has a guanine–adenine base-pair.
An important structural component of RNA that distinguishes it from DNA is the presence of a hydroxyl group at the 2' position of the ribose sugar. The presence of this functional group causes the helix to mostly take the A-form geometry, although in single strand dinucleotide contexts, RNA can rarely also adopt the B-form most commonly observed in DNA. The A-form geometry results in a very deep and narrow major groove and a shallow and wide minor groove. A second consequence of the presence of the 2'-hydroxyl group is that in conformationally flexible regions of an RNA molecule (that is, not involved in formation of a double helix), it can chemically attack the adjacent phosphodiester bond to cleave the backbone.
Structure of a fragment of an RNA, showing a guanosyl subunit.
RNA is transcribed with only four bases (adenine, cytosine, guanine and uracil), but these bases and attached sugars can be modified in numerous ways as the RNAs mature. Pseudouridine (Ψ), in which the linkage between uracil and ribose is changed from a C–N bond to a C–C bond, and ribothymidine (T) are found in various places (the most notable ones being in the TΨC loop of tRNA). Another notable modified base is hypoxanthine, a deaminated adenine base whose nucleoside is called inosine (I). Inosine plays a key role in the wobble hypothesis of the genetic code.
There are more than 100 other naturally occurring modified nucleosides. The greatest structural diversity of modifications can be found in tRNA, while pseudouridine and nucleosides with 2'-O-methylribose often present in rRNA are the most common. The specific roles of many of these modifications in RNA are not fully understood. However, it is notable that, in ribosomal RNA, many of the post-transcriptional modifications occur in highly functional regions, such as the peptidyl transferase center and the subunit interface, implying that they are important for normal function.
The functional form of single-stranded RNA molecules, just like proteins, frequently requires a specific tertiary structure. The scaffold for this structure is provided by secondary structural elements that are hydrogen bonds within the molecule. This leads to several recognizable "domains" of secondary structure like hairpin loops, bulges, and internal loops. Since RNA is charged, metal ions such as Mg2+ are needed to stabilise many secondary and tertiary structures.
The naturally occurring enantiomer of RNA is D-RNA composed of D-ribonucleotides. All chirality centers are located in the D-ribose. By the use of L-ribose or rather L-ribonucleotides, L-RNA can be synthesized. L-RNA is much more stable against degradation by RNase.
Like other structured biopolymers such as proteins, one can define topology of a folded RNA molecule. This is often done based on arrangement of intra-chain contacts within a folded RNA, termed as circuit topology.
Synthesis of RNA is usually catalyzed by an enzyme — RNA polymerase — using DNA as a template, a process known as transcription. Initiation of transcription begins with the binding of the enzyme to a promoter sequence in the DNA (usually found "upstream" of a gene). The DNA double helix is unwound by the helicase activity of the enzyme. The enzyme then progresses along the template strand in the 3’ to 5’ direction, synthesizing a complementary RNA molecule with elongation occurring in the 5’ to 3’ direction. The DNA sequence also dictates where termination of RNA synthesis will occur.
There are also a number of RNA-dependent RNA polymerases that use RNA as their template for synthesis of a new strand of RNA. For instance, a number of RNA viruses (such as poliovirus) use this type of enzyme to replicate their genetic material. Also, RNA-dependent RNA polymerase is part of the RNA interference pathway in many organisms.
Messenger RNA (mRNA) is the RNA that carries information from DNA to the ribosome, the sites of protein synthesis (translation) in the cell. The coding sequence of the mRNA determines the amino acid sequence in the protein that is produced. However, many RNAs do not code for protein (about 97% of the transcriptional output is non-protein-coding in eukaryotes).
These so-called non-coding RNAs (“ncRNA”) can be encoded by their own genes (RNA genes), but can also derive from mRNA introns. The most prominent examples of non-coding RNAs are transfer RNA (tRNA) and ribosomal RNA (rRNA), both of which are involved in the process of translation. There are also non-coding RNAs involved in gene regulation,RNA processing and other roles. Certain RNAs are able to catalyse chemical reactions such as cutting and ligating other RNA molecules, and the catalysis of peptide bond formation in the ribosome; these are known as ribozymes.
Messenger RNA (mRNA) carries information about a protein sequence to the ribosomes, the protein synthesis factories in the cell. It is coded so that every three nucleotides (a codon) corresponds to one amino acid. In eukaryotic cells, once precursor mRNA (pre-mRNA) has been transcribed from DNA, it is processed to mature mRNA. This removes its introns—non-coding sections of the pre-mRNA. The mRNA is then exported from the nucleus to the cytoplasm, where it is bound to ribosomes and translated into its corresponding protein form with the help of tRNA. In prokaryotic cells, which do not have nucleus and cytoplasm compartments, mRNA can bind to ribosomes while it is being transcribed from DNA. After a certain amount of time, the message degrades into its component nucleotides with the assistance of ribonucleases.
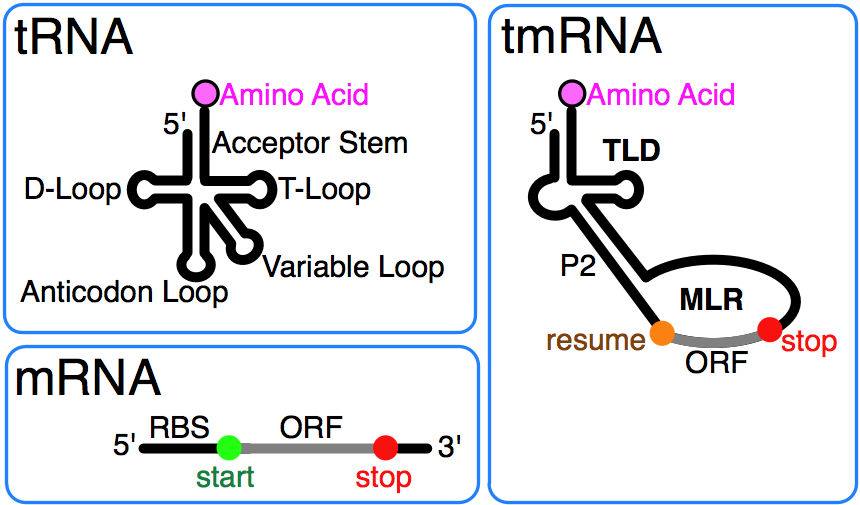
Transfer RNA (tRNA) is a small RNA chain of about 80 nucleotidesthat transfers a specific amino acid to a growing polypeptide chain at the ribosomal site of protein synthesis during translation. It has sites for amino acid attachment and an anticodon region for codon recognition that binds to a specific sequence on the messenger RNA chain through hydrogen bonding.
Ribosomal RNA (rRNA) is the catalytic component of the ribosomes. Eukaryotic ribosomes contain four different rRNA molecules: 18S, 5.8S, 28S and 5S rRNA. Three of the rRNA molecules are synthesized in the nucleolus, and one is synthesized elsewhere. In the cytoplasm, ribosomal RNA and protein combine to form a nucleoprotein called a ribosome. The ribosome binds mRNA and carries out protein synthesis. Several ribosomes may be attached to a single mRNA at any time. Nearly all the RNA found in a typical eukaryotic cell is rRNA.
Transfer-messenger RNA (tmRNA) is found in many bacteria and plastids. It tags proteins encoded by mRNAs that lack stop codons for degradation and prevents the ribosome from stalling.
The earliest known regulators of gene expression were proteins known as repressors and activators, regulators with specific short binding sites within enhancer regions near the genes to be regulated. More recently, RNAs have been found to regulate genes as well. There are several kinds of RNA-dependent processes in eukaryotes regulating the expression of genes at various points, such as RNAi repressing genes post-transcriptionally, long non-coding RNAs shutting down blocks of chromatinepigenetically, and enhancer RNAs inducing increased gene expression. In addition to these mechanisms in eukaryotes, both bacteria and archaea have been found to use regulatory RNAs extensively. Bacterial small RNA and the CRISPR system are examples of such prokaryotic regulatory RNA systems. Fire and Mello were awarded the 2006 Nobel Prize in Physiology or Medicine for discovering microRNAs (miRNAs), specific short RNA molecules that can base-pair with mRNAs.
Post-transcriptional expression levels of many genes can be controlled by RNA interference, in which miRNAs, specific short RNA molecules, pair with mRNA regions and target them for degradation. This antisense-based process involves steps that first process the RNA so that it can base-pair with a region of its target mRNAs. Once the base pairing occurs, other proteins direct the mRNA to be destroyed by nucleases. Fire and Mello were awarded the 2006 Nobel Prize in Physiology or Medicine for this discovery.
Next to be linked to regulation were Xist and other long noncoding RNAs associated with X chromosome inactivation. Their roles, at first mysterious, were shown by Jeannie T. Lee and others to be the silencing of blocks of chromatin via recruitment of Polycomb complex so that messenger RNA could not be transcribed from them. Additional lncRNAs, currently defined as RNAs of more than 200 base pairs that do not appear to have coding potential, have been found associated with regulation of stem cellpluripotency and cell division.
The third major group of regulatory RNAs is called enhancer RNAs. It is not clear at present whether they are a unique category of RNAs of various lengths or constitute a distinct subset of lncRNAs. In any case, they are transcribed from enhancers, which are known regulatory sites in the DNA near genes they regulate. They up-regulate the transcription of the gene(s) under control of the enhancer from which they are transcribed.
At first, regulatory RNA was thought to be a eukaryotic phenomenon, a part of the explanation for why so much more transcription in higher organisms was seen than had been predicted. But as soon as researchers began to look for possible RNA regulators in bacteria, they turned up there as well, termed as small RNA (sRNA). Currently, the ubiquitous nature of systems of RNA regulation of genes has been discussed as support for the RNA World theory. Bacterial small RNAs generally act via antisense pairing with mRNA to down-regulate its translation, either by affecting stability or affecting cis-binding ability. Riboswitches have also been discovered. They are cis-acting regulatory RNA sequences acting allosterically. They change shape when they bind metabolites so that they gain or lose the ability to bind chromatin to regulate expression of genes.
Archaea also have systems of regulatory RNA. The CRISPR system, recently being used to edit DNA in situ, acts via regulatory RNAs in archaea and bacteria to provide protection against virus invaders.
Uridine to pseudouridine is a common RNA modification.
Many RNAs are involved in modifying other RNAs. Introns are spliced out of pre-mRNA by spliceosomes, which contain several small nuclear RNAs (snRNA), or the introns can be ribozymes that are spliced by themselves. RNA can also be altered by having its nucleotides modified to nucleotides other than A,C,G and U. In eukaryotes, modifications of RNA nucleotides are in general directed by small nucleolar RNAs (snoRNA; 60–300 nt), found in the nucleolus and cajal bodies. snoRNAs associate with enzymes and guide them to a spot on an RNA by basepairing to that RNA. These enzymes then perform the nucleotide modification. rRNAs and tRNAs are extensively modified, but snRNAs and mRNAs can also be the target of base modification. RNA can also be methylated.
Like DNA, RNA can carry genetic information. RNA viruses have genomes composed of RNA that encodes a number of proteins. The viral genome is replicated by some of those proteins, while other proteins protect the genome as the virus particle moves to a new host cell. Viroids are another group of pathogens, but they consist only of RNA, do not encode any protein and are replicated by a host plant cell's polymerase.
Reverse transcribing viruses replicate their genomes by reverse transcribing DNA copies from their RNA; these DNA copies are then transcribed to new RNA. Retrotransposons also spread by copying DNA and RNA from one another, and telomerase contains an RNA that is used as template for building the ends of eukaryotic chromosomes.
Double-stranded RNA (dsRNA) is RNA with two complementary strands, similar to the DNA found in all cells, but with the replacement of thymine by uracil. dsRNA forms the genetic material of some viruses (double-stranded RNA viruses). Double-stranded RNA, such as viral RNA or siRNA, can trigger RNA interference in eukaryotes, as well as interferon response in vertebrates.
In the late 1970s, it was shown that there is a single stranded covalently closed, i.e. circular form of RNA expressed throughout the animal and plant kingdom (see circRNA). circRNAs are thought to arise via a "back-splice" reaction where the spliceosome joins a downstream donor to an upstream acceptor splice site. So far the function of circRNAs is largely unknown, although for few examples a microRNA sponging activity has been demonstrated.
Research on RNA has led to many important biological discoveries and numerous Nobel Prizes. Nucleic acids were discovered in 1868 by Friedrich Miescher, who called the material 'nuclein' since it was found in the nucleus. It was later discovered that prokaryotic cells, which do not have a nucleus, also contain nucleic acids. The role of RNA in protein synthesis was suspected already in 1939. Severo Ochoa won the 1959 Nobel Prize in Medicine (shared with Arthur Kornberg) after he discovered an enzyme that can synthesize RNA in the laboratory. However, the enzyme discovered by Ochoa (polynucleotide phosphorylase) was later shown to be responsible for RNA degradation, not RNA synthesis. In 1956 Alex Rich and David Davies hybridized two separate strands of RNA to form the first crystal of RNA whose structure could be determined by X-ray crystallography.
In the early 1970s, retroviruses and reverse transcriptase were discovered, showing for the first time that enzymes could copy RNA into DNA (the opposite of the usual route for transmission of genetic information). For this work, David Baltimore,Renato Dulbecco and Howard Temin were awarded a Nobel Prize in 1975. In 1976, Walter Fiers and his team determined the first complete nucleotide sequence of an RNA virus genome, that of bacteriophage MS2.
In 1977, introns and RNA splicing were discovered in both mammalian viruses and in cellular genes, resulting in a 1993 Nobel to Philip Sharp and Richard Roberts. Catalytic RNA molecules (ribozymes) were discovered in the early 1980s, leading to a 1989 Nobel award to Thomas Cech and Sidney Altman. In 1990, it was found in Petunia that introduced genes can silence similar genes of the plant's own, now known to be a result of RNA interference.
At about the same time, 22 nt long RNAs, now called microRNAs, were found to have a role in the development of C. elegans. Studies on RNA interference gleaned a Nobel Prize for Andrew Fire and Craig Mello in 2006, and another Nobel was awarded for studies on the transcription of RNA to Roger Kornberg in the same year. The discovery of gene regulatory RNAs has led to attempts to develop drugs made of RNA, such as siRNA, to silence genes. Adding to the Nobel prizes awarded for research on RNA in 2009 it was awarded for the elucidation of the atomic structure of the ribosome to Venki Ramakrishnan, Tom Steitz, and Ada Yonath.
Relevance for prebiotic chemistry and abiogenesis
Relevance for prebiotic chemistry and abiogenesis (W)
In 1968, Carl Woese hypothesized that RNA might be catalytic and suggested that the earliest forms of life (self-replicating molecules) could have relied on RNA both to carry genetic information and to catalyze biochemical reactions—an RNA world.
RNA is a single-stranded nucleic acid polymer of the four nucleotides A, C, G, and U joined through a backbone of alternating phosphate and ribosesugar residues. It is the first intermediate in converting the information from DNA into proteins essential for the working of a cell. Some RNAs also serve direct roles in cellular metabolism. RNA is made by copying the base sequence of a section of double-stranded DNA, called a gene, into a piece of single-stranded nucleic acid. This process, called transcription (see belowRNA metabolism), is catalyzed by an enzyme called RNA polymerase.
Whereas DNA provides the genetic information for the cell and is inherently quite stable, RNA has many roles and is much more reactive chemically. RNA is sensitive to oxidizing agents such as periodate that lead to opening of the 3′-terminal ribose ring. The 2′-hydroxyl group on the ribose ring is a major cause of instability in RNA, because the presence of alkali leads to rapid cleavage of the phosphodiester bond linking ribose and phosphate groups. In general, this instability is not a significant problem for the cell, because RNA is constantly being synthesized and degraded.
Interactions between the nitrogen-containing bases differ in DNA and RNA. In DNA, which is usually double-stranded, the bases in one strand pair with complementary bases in a second DNA strand. In RNA, which is usually single-stranded, the bases pair with other bases within the same molecule, leading to complex three-dimensional structures. Occasionally, intermolecular RNA/RNA duplexes do form, but they form a right-handed A-type helix rather than the B-type DNA helix. Depending on the amount of salt present, either 11 or 12 base pairs are found in each turn of the helix. Helices between RNA and DNA molecules also form; these adopt the A-type conformation and are more stable than either RNA/RNA or DNA/DNA duplexes. Such hybrid duplexes are important species in biology, being formed when RNA polymerase transcribes DNA into mRNA for protein synthesis and when reverse transcriptase copies a viral RNA genome such as that of the human immunodeficiency virus (HIV).
Single-stranded RNAs are flexible molecules that form a variety of structures through internal base pairing and additional non-base pair interactions. They can form hairpin loops such as those found in transfer RNA (tRNA), as well as longer-range interactions involving both the bases and the phosphate residues of two or more nucleotides. This leads to compact three-dimensional structures. Most of these structures have been inferred from biochemical data, since few crystallographic images are available for RNA molecules. In some types of RNA, a large number of bases are modified after the RNA is transcribed. More than 90 different modifications have been documented, including extensive methylations and a wide variety of substitutions around the ring. In some cases these modifications are known to affect structure and are essential for function.
Messenger RNA (mRNA) delivers the information encoded in one or more genes from the DNA to the ribosome, a specialized structure, or organelle, where that information is decoded into a protein. In prokaryotes, mRNAs contain an exact transcribed copy of the original DNA sequence with a terminal 5′-triphosphate group and a 3′-hydroxyl residue. In eukaryotes the mRNA molecules are more elaborate. The 5′-triphosphate residue is further esterified, forming a structure called a cap. At the 3′ ends, eukaryotic mRNAs typically contain long runs of adenosine residues (polyA) that are not encoded in the DNA but are added enzymatically after transcription. Eukaryotic mRNA molecules are usually composed of small segments of the original gene and are generated by a process of cleavage and rejoining from an original precursor RNA (pre-mRNA) molecule, which is an exact copy of the gene (as described in the section Splicing). In general, prokaryotic mRNAs are degraded very rapidly, whereas the cap structure and the polyA tail of eukaryotic mRNAs greatly enhance their stability.
Ribosomal RNA (rRNA) molecules are the structural components of the ribosome. The rRNAs form extensive secondary structures and play an active role in recognizing conserved portions of mRNAs and tRNAs. They also assist with the catalysis of protein synthesis. In the prokaryote E. coli, seven copies of the rRNA genes synthesize about 15,000 ribosomes per cell. In eukaryotes the numbers are much larger. Anywhere from 50 to 5,000 sets of rRNA genes and as many as 10 million ribosomes may be present in a single cell. In eukaryotes these rRNA genes are looped out of the main chromosomal fibres and coalesce in the presence of proteins to form an organelle called the nucleolus. The nucleolus is where the rRNA genes are transcribed and the early assembly of ribosomes takes place.
Transfer RNA (tRNA) carries individual amino acids into the ribosome for assembly into the growing polypeptide chain. The tRNA molecules contain 70 to 80 nucleotides and fold into a characteristic cloverleaf structure. Specialized tRNAs exist for each of the 20 amino acids needed for protein synthesis, and in many cases more than one tRNA for each amino acid is present. The nucleotide sequence is converted into a protein sequence by translating each three-base sequence (called a codon) with a specific protein. The 61 codons used to code amino acids can be read by many fewer than 61 distinct tRNAs (as described in the section Translation). In E. coli a total of 40 different tRNAs are used to translate the 61 codons. The amino acids are loaded onto the tRNAs by specialized enzymes called aminoacyl tRNA synthetases, usually with one synthetase for each amino acid. However, in some organisms, less than the full complement of 20 synthetases are required because some amino acids, such as glutamine and asparagine, can be synthesized on their respective tRNAs. All tRNAs adopt similar structures because they all have to interact with the same sites on the ribosome.
Not all catalysis within the cell is carried out exclusively by proteins. Thomas Cech and Sidney Altman, jointly awarded a Nobel Prize in 1989, discovered that certain RNAs, now known as ribozymes, showed enzymatic activity. Cech showed that a noncoding sequence (intron) in the small subunit rRNA of protozoans, which had to be removed before the rRNA was functional, can excise itself from a much longer precursor RNA molecule and rejoin the two ends in an autocatalytic reaction. Altman showed that the RNA component of an RNA protein complex called ribonuclease P can cleave a precursor tRNA to generate a mature tRNA. In addition to self-splicing RNAs similar to the one discovered by Cech, artificial RNAs have been made that show a variety of catalytic reactions. It is now widely held that there was a stage during evolution when only RNA catalyzed and stored genetic information. This period, sometimes called “the RNA world,” is believed to have preceded the function of DNA as genetic material.
Most antisense RNAs are synthetically modified derivatives of RNA or DNA with potential therapeutic value. In nature, antisense RNAs contain sequences that are the complement of the normal coding sequences found in mRNAs (also called sense RNAs). Like mRNAs, antisense RNAs are single-stranded, but they cannot be translated into protein. They can inactivate their complementary mRNA by forming a double-stranded structure that blocks the translation of the base sequence. Artificially introducing antisense RNAs into cells selectively inactivates genes by interfering with normal RNA metabolism.
Many viruses use RNA for their genetic material. This is most prevalent among eukaryotic viruses, but a few prokaryotic RNA viruses are also known. Some common examples include poliovirus, human immunodeficiency virus (HIV), and influenza virus, all of which affect humans, and tobacco mosaic virus, which infects plants. In some viruses the entire genetic material is encoded in a single RNA molecule, while in the segmented RNA viruses several RNA molecules may be present. Many RNA viruses such as HIV use a specialized enzyme called reverse transcriptase that permits replication of the virus through a DNA intermediate. In some cases this DNA intermediate becomes integrated into the host chromosome during infection; the virus then exists in a dormant state and effectively evades the host immune system.
Many other small RNA molecules with specialized functions are present in cells. For example, small nuclear RNAs (snRNAs) are involved in RNA splicing (see below), and other small RNAs that form part of the enzymes telomerase or ribonuclease P are part of ribonucleoprotein particles. The RNA component of telomerase contains a short sequence that serves as a template for the addition of small strings of oligonucleotides at the ends of eukaryotic chromosomes. Other RNA molecules serve as guide RNAs for editing, or they are complementary to small sections of rRNA and either direct the positions at which methyl groups need to be added or mark U residues for conversion to the isomer pseudouridine.
Following synthesis by transcription, most RNA molecules are processed before reaching their final form. Many rRNA molecules are cleaved from much larger transcripts and may also be methylated or enzymatically modified. In addition, tRNAs are usually formed as longer precursor molecules that are cleaved by ribonuclease P to generate the mature 5′ end and often have extra residues added to their 3′ end to form the sequence CCA. The hydroxyl group on the ribose ring of the terminal A of the 3′-CCA sequence acts as the amino acid acceptor necessary for the function of RNA in protein building.
In prokaryotes the protein coding sequence occupies one continuous linear segment of DNA. However, in eukaryotic genes the coding sequences are frequently “split” in the genome—a discovery reached independently in the 1970s by Richard J. Roberts (the author of this article) and Phillip A. Sharp, whose work won them a Nobel Prize in 1993. The segments of DNA or RNA coding for protein are called exons, and the noncoding regions separating the exons are called introns. Following transcription, these coding sequences must be joined together before the mRNAs can function. The process of removal of the introns and subsequent rejoining of the exons is called RNA splicing. Each intron is removed in a separate series of reactions by a complicated piece of enzymatic machinery called a spliceosome. This machinery consists of a number of small nuclear ribonucleoprotein particles (snRNPs) that contain small nuclear RNAs (snRNAs).
Some RNA molecules, particularly those in protozoan mitochondria, undergo extensive editing following their initial synthesis. During this editing process, residues are added or deleted by a posttranscriptional mechanism under the influence of guide RNAs. In some cases as much as 40 percent of the final RNA molecule may be derived by this editing process, rather than being coded directly in the genome. Some examples of editing have also been found in mRNA molecules, but these appear much more limited in scope.
RNA provides the link between the genetic information encoded in DNA and the actual workings of the cell. Some RNA molecules such as the rRNAs and the snRNAs (described in the section Types of RNA) become part of complicated ribonucleoprotein structures with specialized roles in the cell. Others such as tRNAs play key roles in protein synthesis, while mRNAs direct the synthesis of proteins by the ribosome. Three distinct phases of RNA metabolism occur. First, selected segments of the genome are copied by transcription to produce the precursor RNAs. Second, these precursors are processed to become functionally mature RNAs ready for use. When these RNAs are mRNAs, they are then used for translation. Third, after use the RNAs are degraded, and the bases are recycled. Thus, transcription is the process where a specific segment of DNA, a gene, is copied into a specific RNA that encodes a single protein or plays a structural or catalytic role. Translation is the decoding of the information within mRNA molecules that takes place on a specialized structure called a ribosome. There are important differences in both transcription and translation between prokaryotic and eukaryotic organisms.
Small segments of DNA are transcribed into RNA by the enzyme RNA polymerase, which achieves this copying in a strictly controlled process. The first step is to recognize a specific sequence on DNA called a promoter that signifies the start of the gene. The two strands of DNA become separated at this point, and RNA polymerase begins copying from a specific point on one strand of the DNA using a ribonucleoside 5′-triphosphate to begin the growing chain. Additional ribonucleoside triphosphates are used as the substrate, and, by cleavage of their high-energy phosphate bond, ribonucleoside monophosphates are incorporated into the growing RNA chain. Each successive ribonucleotide is directed by the complementary base pairing rules of DNA. Thus, a C in DNA directs the incorporation of a G into RNA, G is copied into C, T into A, and A into U. Synthesis continues until a termination signal is reached, at which point the RNA polymerase drops off the DNA, and the RNA molecule is released. In some cases this RNA molecule is the final mRNA. In other cases it is a pre-mRNA and requires further processing before it is ready for translation by the ribosome. Ahead of many genes in prokaryotes, there are signals called “operators” where specialized proteins called repressors bind to the DNA just upstream of the start point of transcription and prevent access to the DNA by RNA polymerase. These repressor proteins thus prevent transcription of the gene by physically blocking the action of the RNA polymerase. Typically, repressors are released from their blocking action when they receive signals from other molecules in the cell indicating that the gene needs to be expressed. Ahead of some prokaryotic genes are signals to which activator proteins bind that positively induce transcription.
Transcription in higher organisms is more complicated. First, the RNA polymerase of eukaryotes is a more complicated enzyme than the relatively simple five-subunit enzyme of prokaryotes. In addition, there are many more accessory factors that help to control the efficiency of the individual promoters. These accessory proteins are called transcription factors and typically respond to signals from within the cell that indicate whether transcription is required. In many human genes, several transcription factors may be needed before transcription can proceed efficiently. A transcription factor can cause either repression or activation of gene expression in eukaryotes.
During transcription, only one strand of the DNA is usually copied. This is called the template strand, and the RNA molecules produced are single-stranded. The DNA strand that would correspond to the mRNA is called the coding or sense strand, and it is not unusual for this to change from one gene to the next. In eukaryotes the initial product of transcription is called a pre-mRNA, which is extensively spliced before the mature mRNA is produced, ready for translation by the ribosome.
The process of translation uses the information present in the nucleotide sequence of mRNA to direct the synthesis of a specific protein for use by the cell. Translation takes place on the ribosomes — complex particles in the cell that contain RNA and protein. In prokaryotes the ribosomes are loaded onto the mRNA while transcription is still ongoing. Near the 5′ end of the mRNA, a short sequence of nucleotides signals the starting point for translation. It contains a few nucleotides called a ribosome binding site, or Shine-Dalgarno sequence. In E. coli the tetranucleotide GAGG is sufficient to serve as a binding site. This typically lies five to eight bases upstream of an initiation codon. The mRNA sequence is read three bases at a time from its 5′ end toward its 3′ end, and one amino acid is added to the growing chain from its respective aminoacyl tRNA, until the complete protein chain is assembled. Translation stops when the ribosome encounters a termination codon, normally UAG, UAA, or UGA. Special release factors associate with the ribosome in response to these codons, and the newly synthesized protein, tRNAs, and mRNA all dissociate. The ribosome then becomes available to interact with another mRNA molecule.
In eukaryotes the essence of protein synthesis is the same, although the ribosomes are more complicated. As with prokaryotic initiation, the signal sequence interacts with the 3′ end of the small subunit rRNA during formation of the initiation complex.
The issue of fidelity is important during protein synthesis, but it is not as crucial as fidelity during replication. One mRNA molecule can be translated repeatedly to give many copies of the protein. When an occasional protein is mistranslated, it usually does not fold properly and is then degraded by the cellular machinery. However, proofreading mechanisms exist within the ribosome to ensure accurate pairing between the codon in the mRNA and the anticodon in the tRNA.
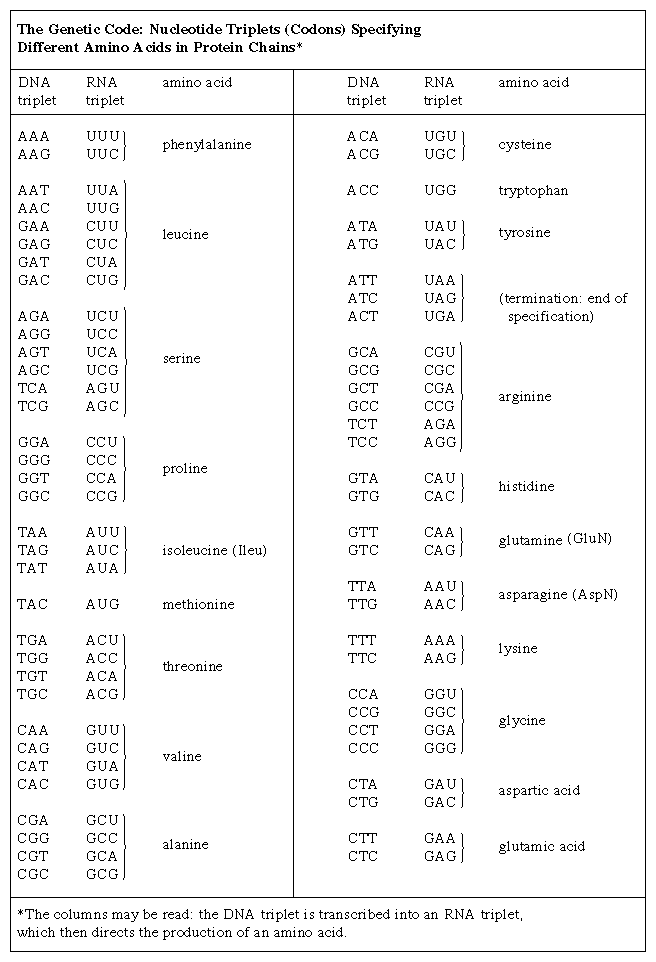
One of the crowning achievements of molecular biology was the elucidation during the 1960s of the genetic code. Principals in this effort were Har G. Khorana and Marshall W. Nirenberg, who shared a Nobel Prize in 1968. Khorana and Nirenberg used artificial templates and protein synthesizing systems in the test tube to determine the coding potential of all 64 possible triplet codons (see the table). The key feature of the genetic code is that the 20 amino acids are encoded by 61 codons. Thus, there is degeneracy in the code such that one amino acid is often specified by more than one codon. In the case of serine and leucine, six codons can be used for each. Among organisms that have been examined in detail, the code appears to be almost universal, from bacteria through archaea to eukaryotes. The known exceptions are found in the mitochondria of humans and many other organisms as well as in some species of bacteria. The structure within the genetic code whereby many amino acids are uniquely coded by the first two bases of the codon strongly suggests that the code has itself evolved from a more primitive code involving 16 dinucleotides. How the individual amino acids became associated with the different codons remains a matter of speculation.
All living creatures today reproduce and evolve using a complex gene-enzyme cycle. If we look at a cell, for example, information encoded in its genes is used to produce functional proteins called enzymes. Some of those enzymes then turn around to make copies of the cell's genes, allowing the cell to reproduce.
Because the gene-enzyme system forms a closed loop, it presents us with a classic chicken or egg conundrum: Which came first, genes or the protein enzymes they code for?
While the details are still not fully worked out, discoveries over the past few decades have lead researchers to a surprising possible solution: What really came first? Genes that act as enzymes!
The RNA World Hypothesis is the idea that before living cells, the genetic code, and the gene/protein cycle ever existed, chains of a chemical called RNA were forming naturally. Once formed, some of these chains were able to function as enzymes, and were even able to evolve by making copies of themselves with slight, accidental modifications.
While there is little doubt that RNA played a crucial role in the early development of life, the complexity of RNA is casting doubt on the idea that RNA was the first truly replicating and evolving chemical system. For this reason, alternatives to the “RNA first” view are also being investigated. Most notable is a proto-RNA hypothesis being studied by the lab of Nicholas Hud, and several metabolism first hypotheses which got their start from the work of Robert Shapiro.
At this time, all serious investigations into the origin of life are being done under the overarching idea that life emerged from chemistry.
📹 The RNA Origin of Life / NOVA PBS Official (VİDEO)
📹 The RNA Origin of Life / NOVA PBS Official (LINK)
RNA may have been the origin of life on Earth. Go on a whirlwind tour of RNA's evolving role through billions of years of evolutionary history.
📹 Origins of Life — Early Life - RNA / The RNA World — Complexity Explorer (VİDEO)
📹 Origins of Life — Early Life - RNA - The RNA World / Complexity Explorer (LINK)
The Origins of the RNA World | Library of Congress
📹 The Origins of the RNA World | Library of Congress




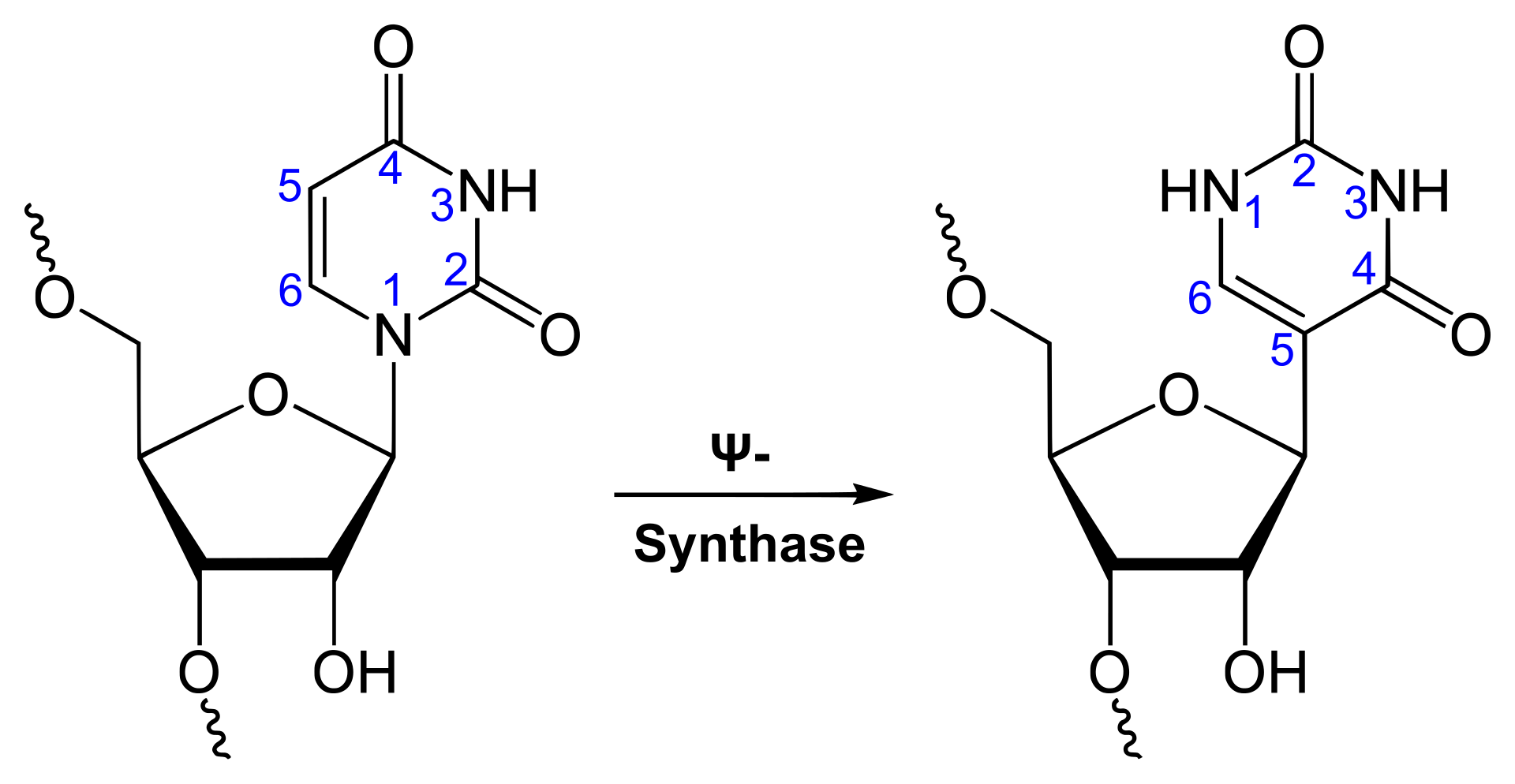



{kind=link}