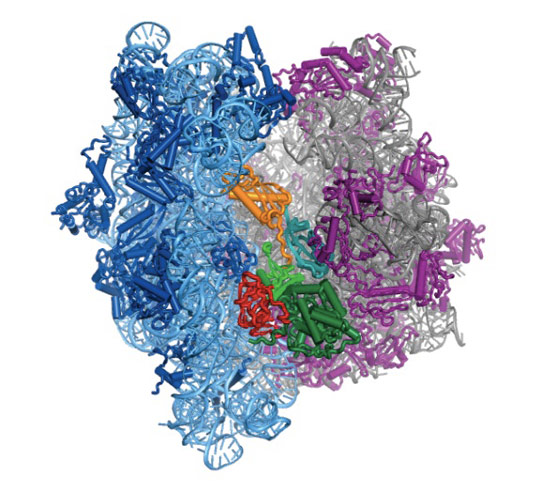
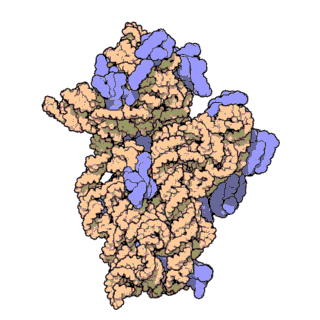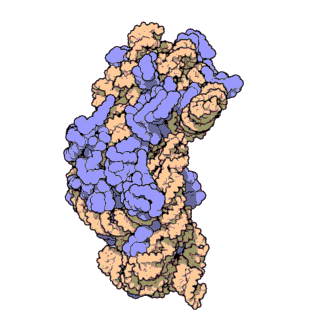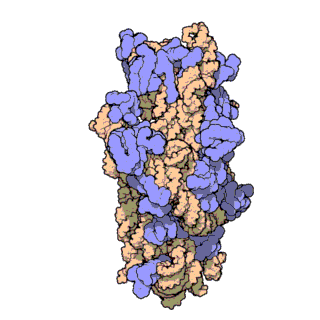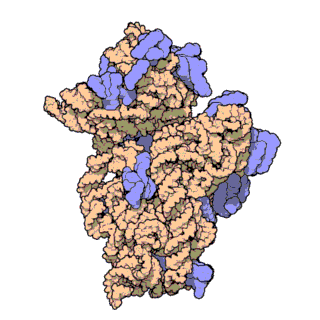
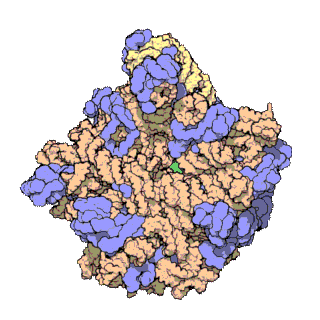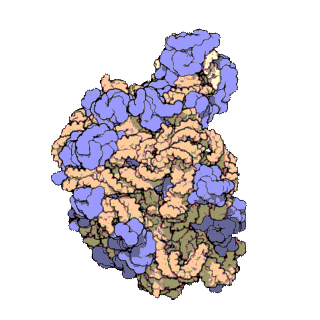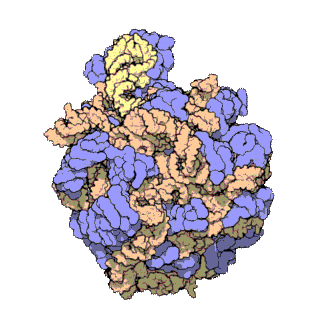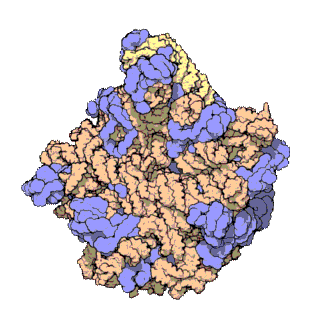
Bakteride protein yapıcı ribozomun yapısal incelemesi uzatma etmeni G denilen bir molekülün (özekteki sarı-yeşil-kırmızı yapı) ribozomun devimini nasıl denetlediğini gösterdi.
Structural studies of the protein-making ribosome in bacteria shed light on how a molecule called elongation factor G (yellow-green-red structure in center) controls its motion.
Here's the kind of equipment required to create that image. This is the endstation of an Advanced Light Source beamline called SIBYLS, or Structurally Integrated Biology for Life Sciences. (LINK)
🛑 ANAHATLAR
ANAHATLAR
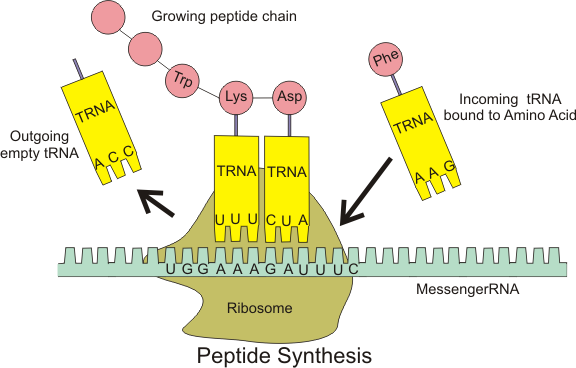
Protein bireşimi ribozomlar aracılığıyla yerine getirilir.
(Bağlantılı kavramlar: Moleküler biolojinin özeksel inağı; gen anlatımı)
mRNA → protein çevrilmesi ribozomlarda yer alır.
tRNA antikodonu mRNA kodonu ile buluşur ve amino asit polipeptide eklenmek üzere ribozoma bırakılır.
Ortalama memeli hücresinde 10 milyon kadar ribozom bulunur.
Ribozomlar hem serbest parçacıklar olarak hem de endoplazmik retikulumun membranlarına bağlanmış olarak bulunur.
Ribozomlar ilk kez 1955’te betimlendi.
🛑 RİBOZOM YAPISI
RİBOZOM YAPISI
Ribozomlar tüm hücrelerde bulunan ve yaklaşık olarak eşit ölçülerde ribozomal RNA (rRNA) ve ribozomal proteinlerden oluşan makromoleküler düzeneklerdir.
Her bir ribozom biri küçük ve biri büyük iki altbirimden oluşur.
Her bir alt-birim bir ya da birçok ribozomal RNA (rRNA) molekülünden ve birçok ribozomal proteinden (RPler ya da r-proteinler) oluşur (ökaryotlarda genellikle üç ya da dört rRNA molekülü ve 40-80 arasında ribozomal protein).
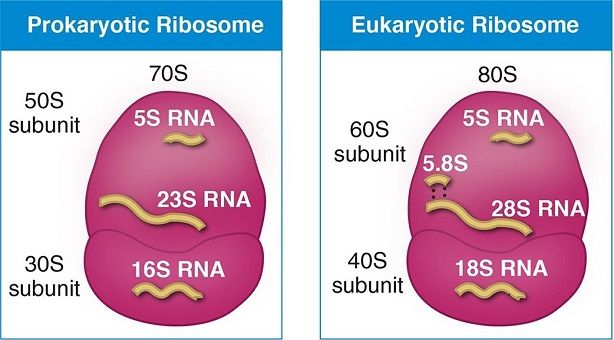
Ribozomun altbirimleri bir santrifüj alanında Svedberg birimlerinde (S) ölçülen çöküm (sedimentasyon) oranlarının terimlerinde belirtilir
Ökaryotların küçük ve büyük ribozom altbirimleri sırasıyla 40S ve 60S olarak belirtilir (prokaryotlarda 30S ve 50S).
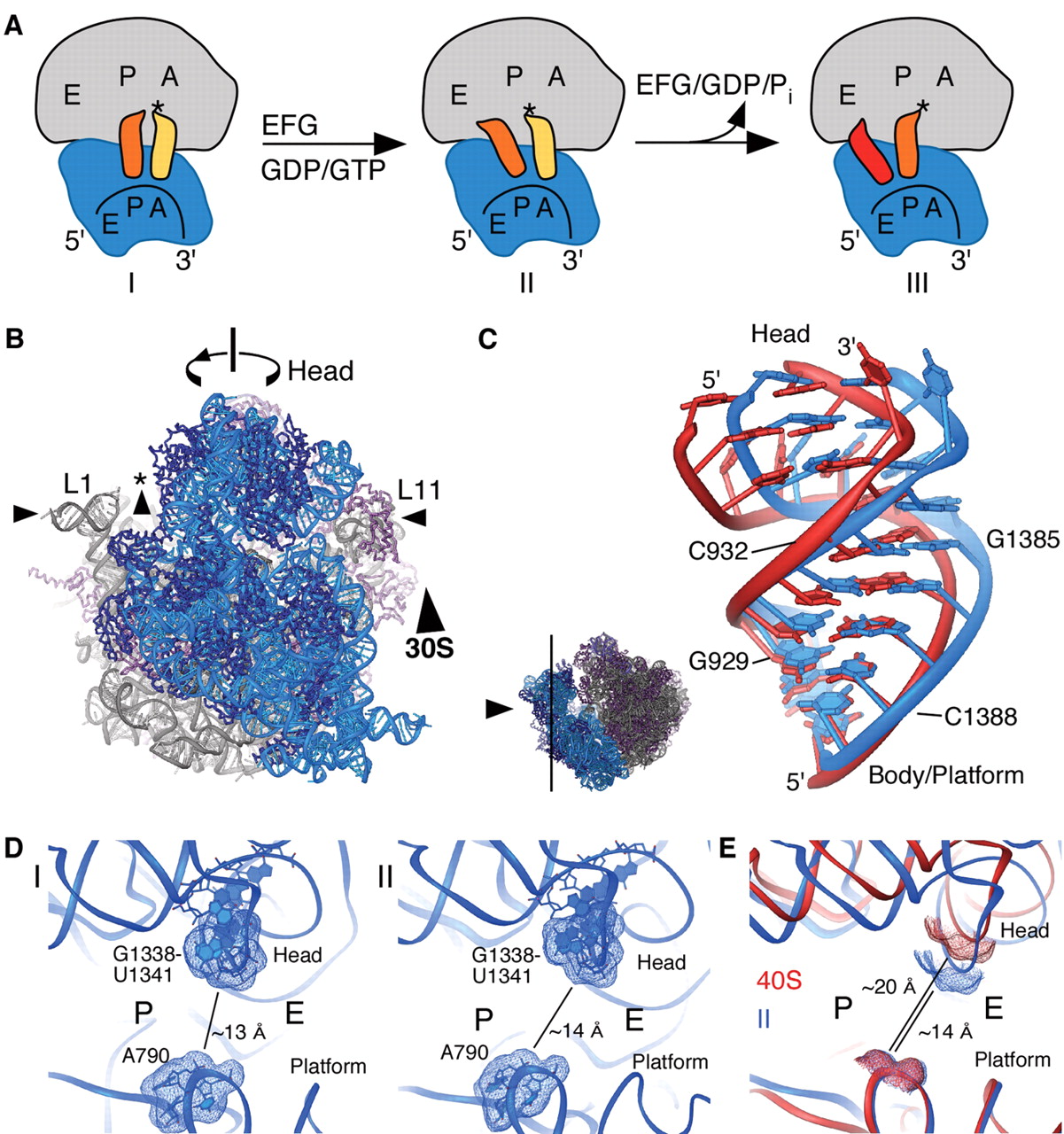
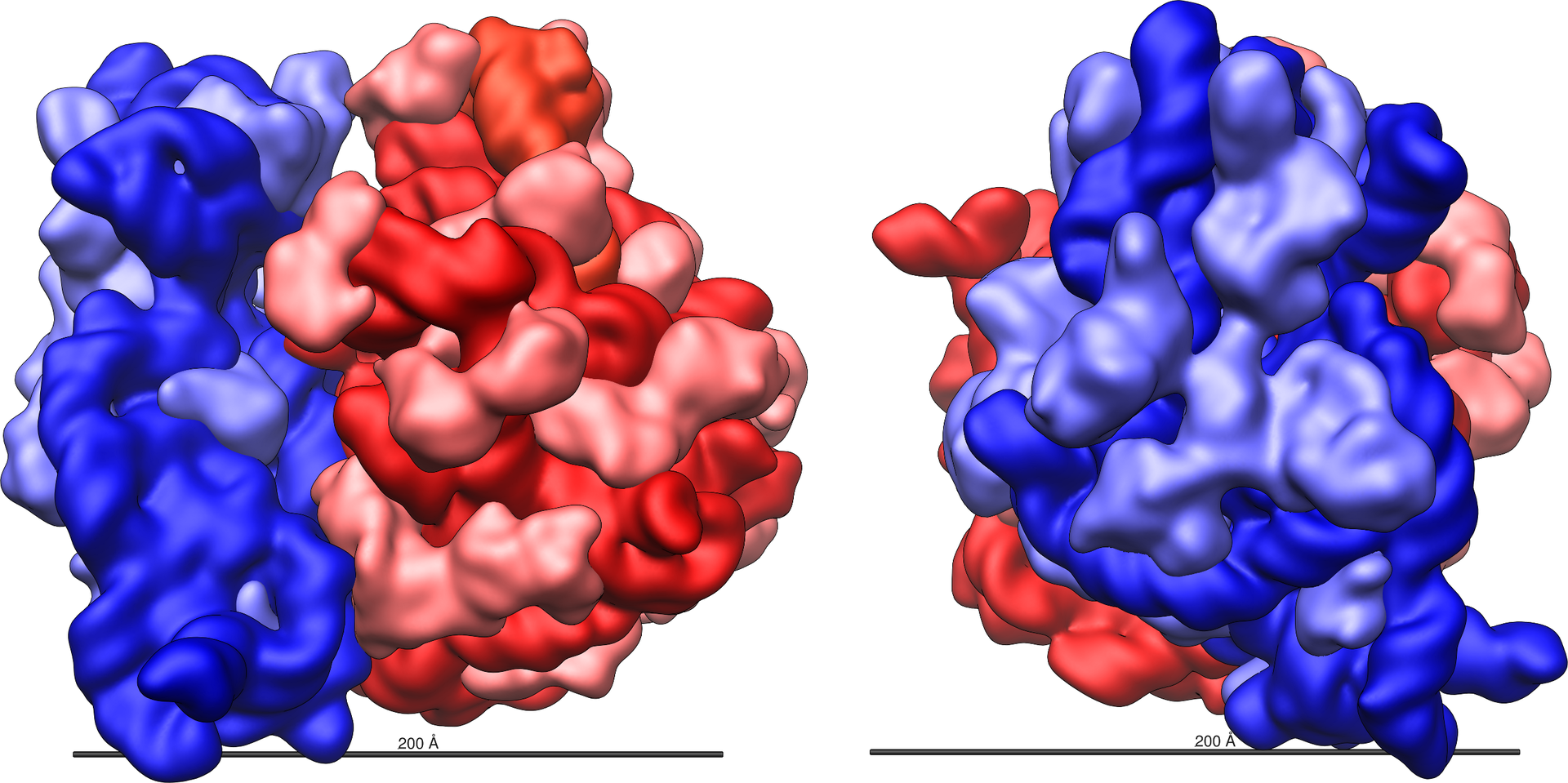
Small-subunit tRNA binding sites and translocation intermediates(LINK)
Small-subunit tRNA binding sites and translocation intermediates.
(A) Schematic steps in the mRNA and tRNA translocation reaction. After peptide bond formation, the pretranslocation complex (I) binds EF-G and shifts to the hybrid state of tRNA binding (complex II). Subsequent GTP hydrolysis by EF-G leads to the posttranslocation state (complex III). [Model abbreviated from (31)]
(B) Model of structural changes in the ribosome that contribute to translocation, viewed as in Fig. 1A. Arrows indicate motions of the small and large subunit proposed to occur during the ratcheting mechanism: ratcheting of the small subunit, rotation of the small-subunit head domain, opening of the tRNA binding groove (asterisk), and lateral movements of the L1 and L11 arms (16, 21, 27).
(C) View of the conformational differences in h28 of 16S rRNA in ribosomes I (blue) and II (red). The direction of view is indicated to the left.
(D) View of the tRNA binding cleft in the 30S subunit, from the perspective of the large subunit. The G1338-U1341 ridge and A790, which separate the P and E sites, are marked. Molecular surfaces of the tRNA binding cleft in ribosome I (left) and ribosome II (right) are indicated. The shortest distance between the van der Waals surfaces is marked in each case.
(E) Superposition of the 30S subunit in ribosome II (blue) with the yeast 40S subunit from the 80S-eEF2-sardorin complex (red) (27, 51). Molecular surfaces and distances are shown as in Fig. 2D.
🛑 RİBOZOM ETKİNLİĞİ
RİBOZOM ETKİNLİĞİ
Ribozom mRNAyı okur ve biolojik protein bireşimini (mRNA çevirisi) yerine getirir.
Ribozomlar amino asitleri iletmen RNA (mRNA) kodonları tarafından belirlenen düzen içinde biraraya getirerek polipeptid bireşimini sağlar.
tRNA moleküllerinin düzeni bir proteinin amino asit dizisini belirler.
rRNA molekülleri amino asitler arasında peptid bağları kurarak proteinleri oluşturan peptidil tranferaz tepkimesine katalizörlük eder (ribozomlar peptidil hidrolisiste de katalizör olarak davranır).
Yeni oluşan proteinler ribozom sitesinden kopar ve kullanım için hücrenin başka yerlerine göç eder.
Making mitochondrial hydrophobic proteins
Mitochondria produce chemical energy for the cell. Human mitochondria have their own specific ribosomes—mitoribosomes, which are distinct from cytoplasmic ribosomes. Mitoribosomes synthesize the mitochondrial membrane proteins that generate the chemical energy. Brown et al. used cryo–electron microscopy to determine the high-resolution structure of the large subunit of the human mitoribosome. The mitoribosome has a number of unique features, including an exit tunnel lined with hydrophobic amino acid residues.
Abstract
Human mitochondrial ribosomes are highly divergent from all other known ribosomes and are specialized to exclusively translate membrane proteins. They are linked with hereditary mitochondrial diseases and are often the unintended targets of various clinically useful antibiotics. Using single-particle cryogenic electron microscopy, we have determined the structure of its large subunit to 3.4 angstrom resolution, revealing 48 proteins, 21 of which are specific to mitochondria. The structure unveils an adaptation of the exit tunnel for hydrophobic nascent peptides, extensive remodeling of the central protuberance, including recruitment of mitochondrial valine transfer RNA (tRNAVal) to play an integral structural role, and changes in the tRNA binding sites related to the unusual characteristics of mitochondrial tRNAs.
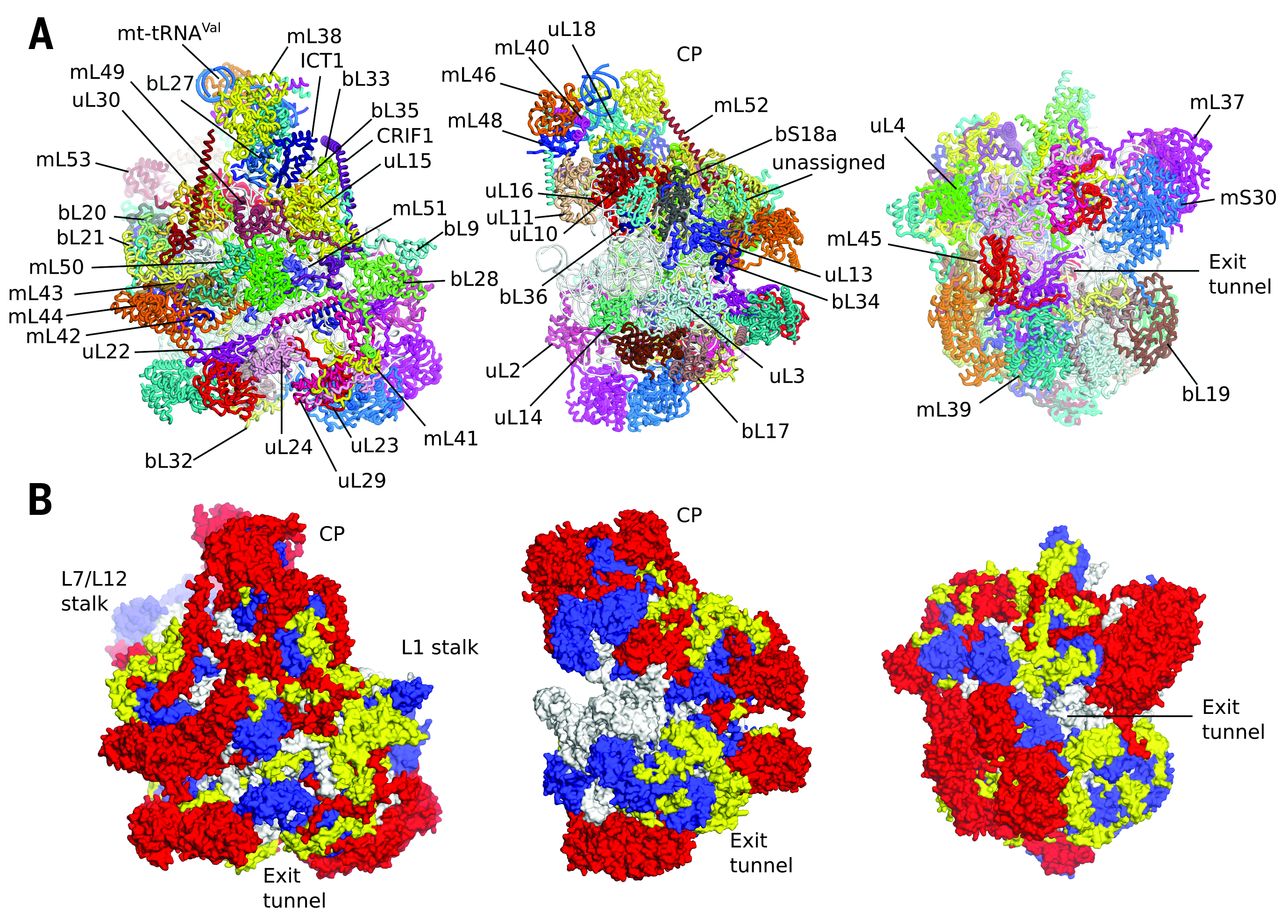
Fig. 1 Overview of human mt-LSU.
(A) Location of proteins in the human mt-LSU, showing (from left to right) solvent-facing, side, and exit tunnel views. (B) Views as in A, proteins conserved with bacteria (blue), extensions of homologous proteins (yellow), and mitochondria-specific proteins (red). rRNA is shown in gray.
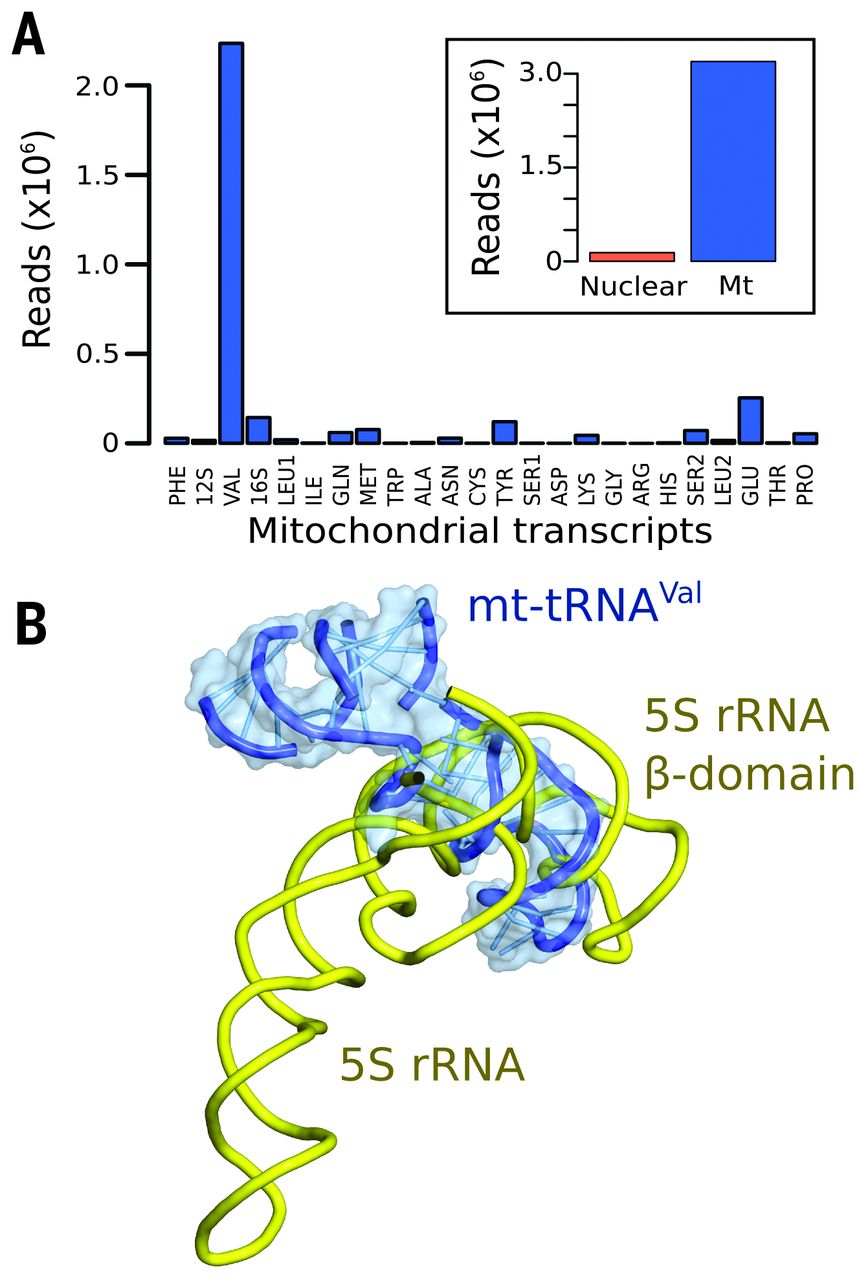
Fig. 2 Mt-tRNAVal is part of the human mitoribosome.
(A) Mapping of rRNA-sequencing reads to total human (inset) and mitochondrial transcripts. (B) The anticodon stem-loop of mt-tRNAVal binds in a similar position to domain β of 5S rRNA..
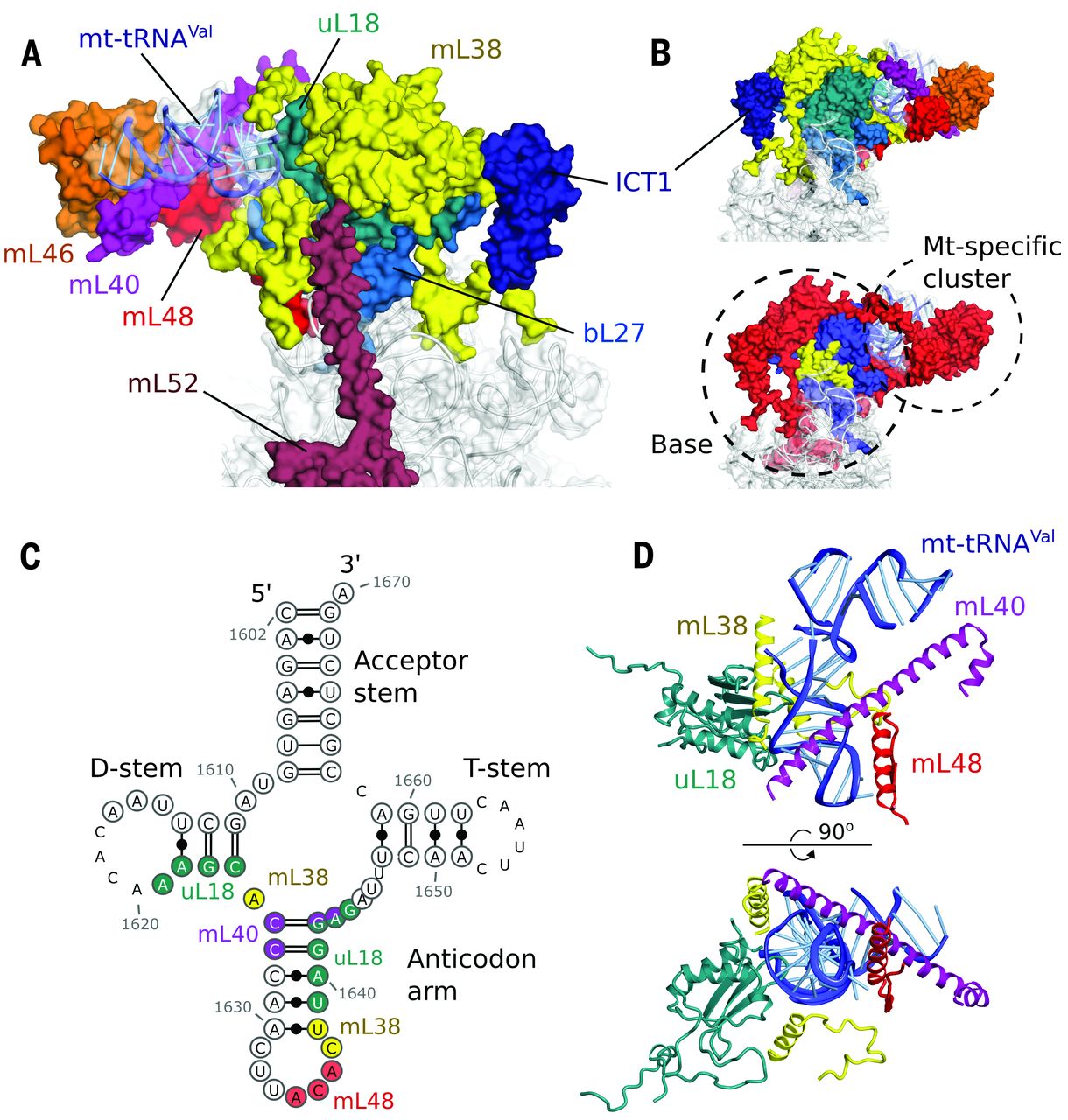
Fig. 3 The central protuberance containing mt-tRNAVal.
(A) Relative locations of proteins and mt-tRNAVal in the central protuberance. (B) View of (A) rotated by 180°, colored by proteins (top) and conservation (bottom) in accordance with Fig. 1. (C) Secondary structure of mt-tRNAVal. Modeled nucleotides are circled, and those interacting with surrounding proteins are colored. (D) The anticodon arm of mt-tRNAVal (blue) interacts extensively with proteins, whereas the acceptor arm is solvent exposed..
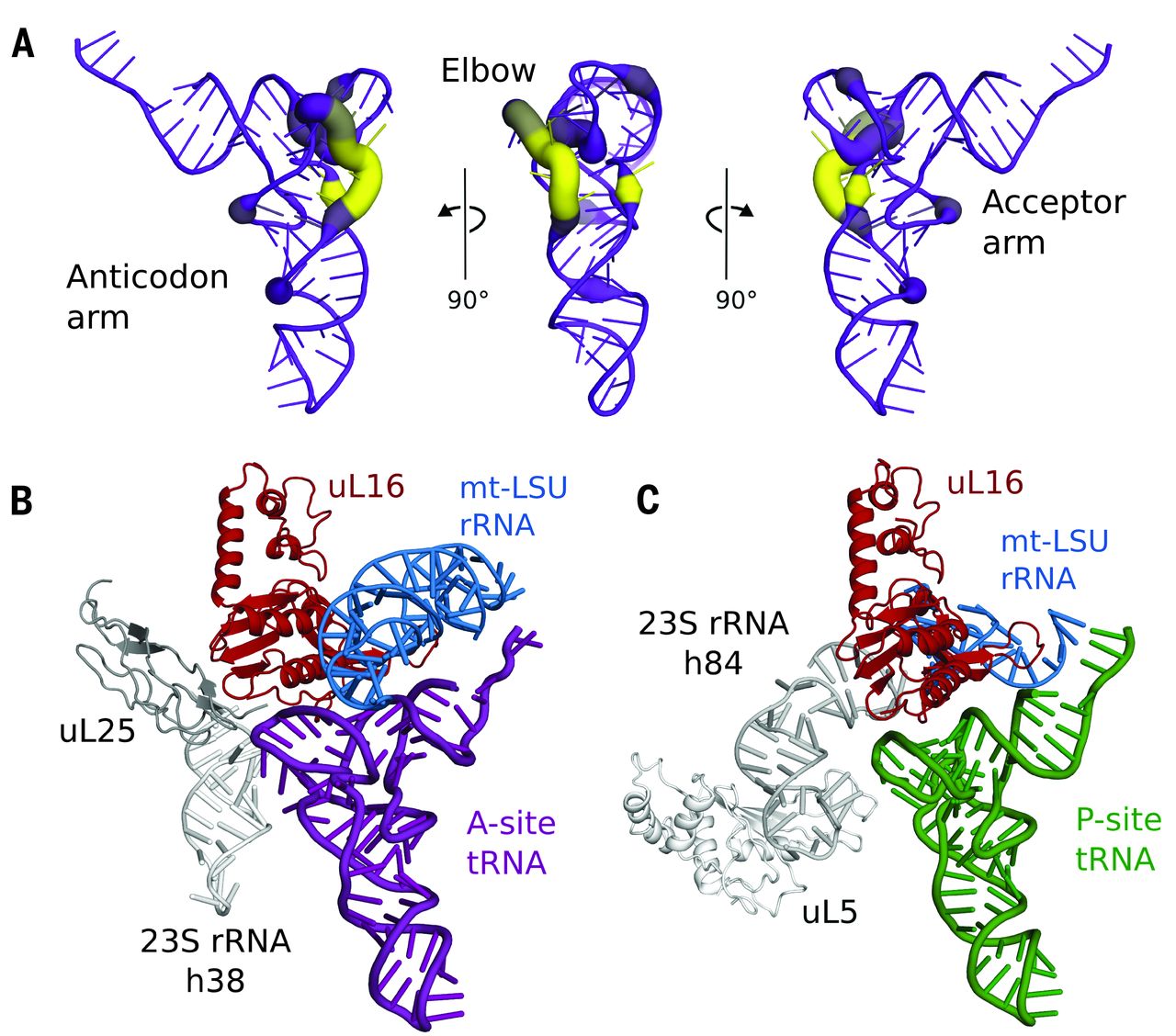
Fig. 4 Coevolution of mt-tRNAs and their binding sites.
(A) Variability in the elbow region of human mt-tRNAs. The deletion of nucleotides relative to a bacterial tRNA (PDB ID: 2WDI) is shown by line color and thickness, with yellow and thick lines indicating most frequently deleted. (B) Modeling a bacterial A-site tRNA (purple) reveals that uL25 and 23S rRNA h38 (both gray) that stabilize the tRNA elbow region are deleted compared with bacterial ribosomes. (C) Similarly, uL5 and 23S rRNA h84 (both gray) that stabilize the elbow region of P-site tRNA (green) are deleted, but elements that bind the anticodon arm are conserved..
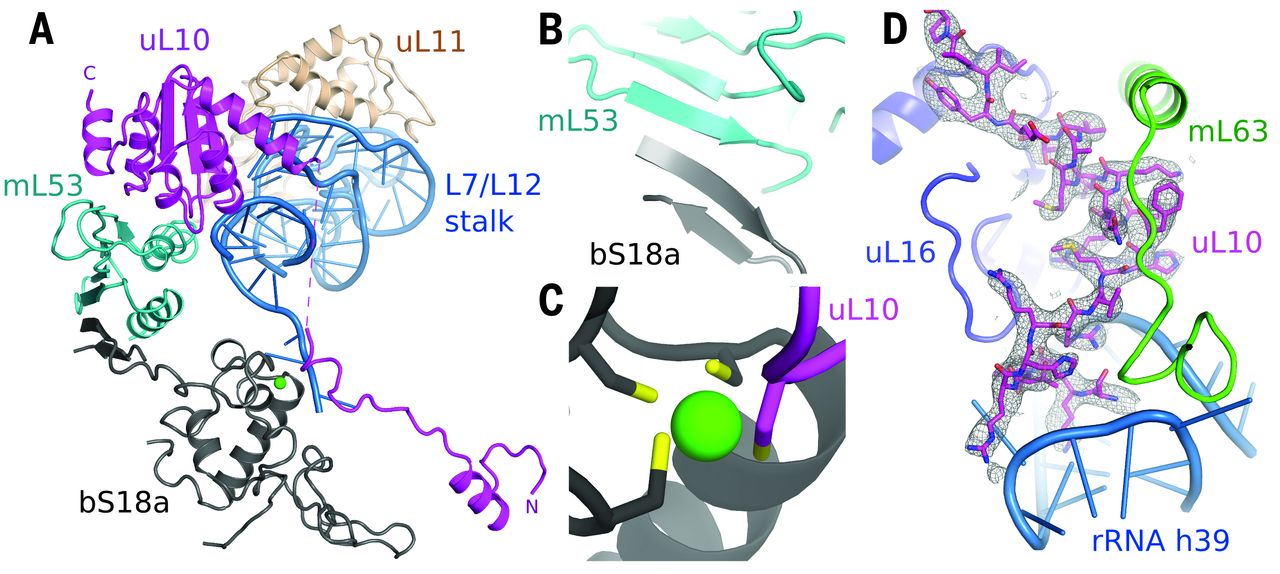
Fig. 5 Remodeling of the L7/L12 stalk.
(A) Overview of new elements at the L7/L12 stalk. (B) bS18a forms a shared β sheet with mL53 to connect the stalk to the body of the mitoribosome. (C) The novel N-terminal extension of uL10 contributes a cysteine residue to a shared zinc-binding motif with bS18a. (D) Density for the N-terminal extension of uL10 that is highly coordinated to the body of the ribosome.
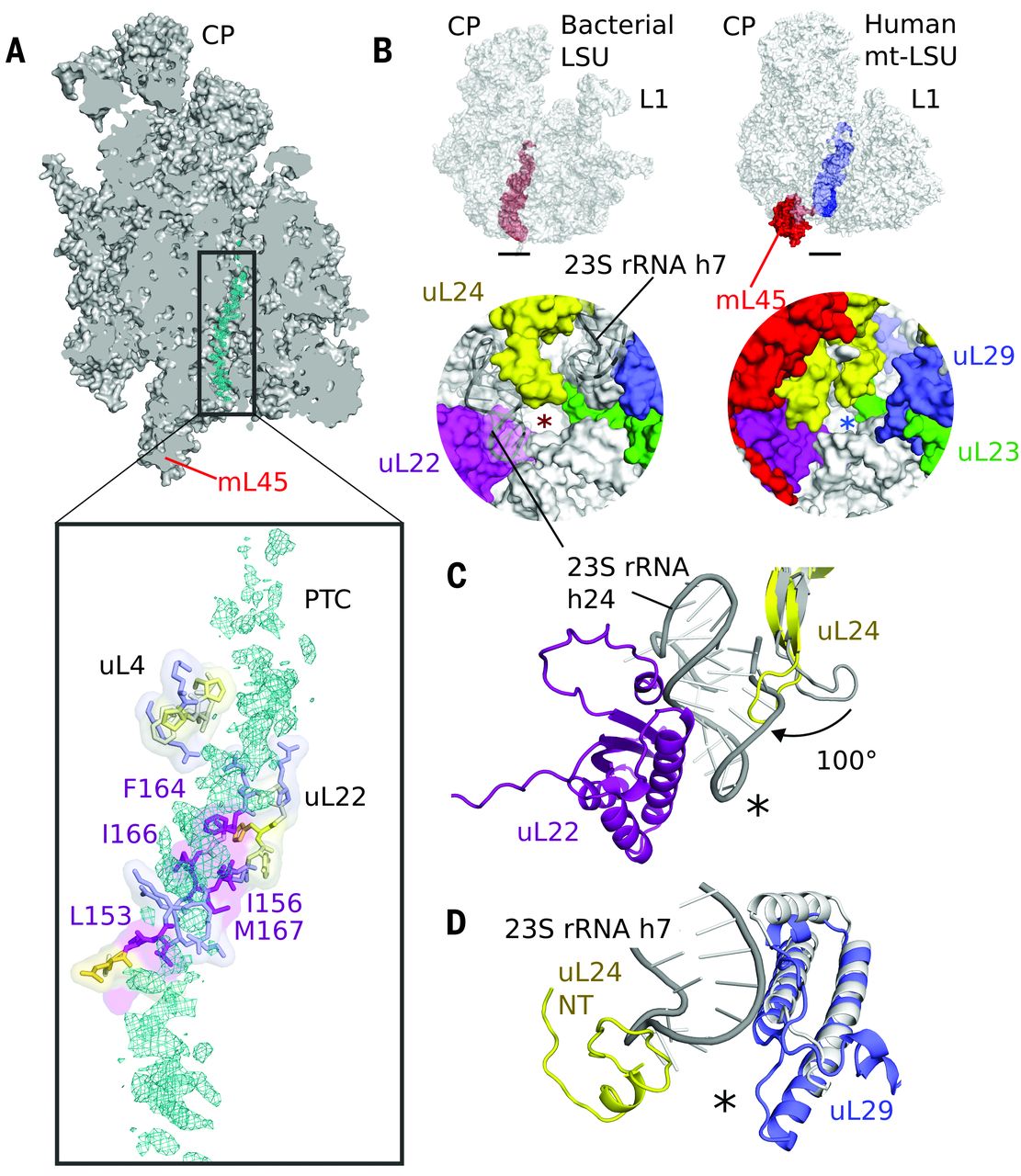
Fig. 6 The exit tunnel.
(A) Slice through the mt-LSU showing nascent chain density (cyan) in the exit tunnel. The nascent polypeptide interacts with a β hairpin of uL22 enriched with hydrophobic residues. (B) The exit tunnel in bacteria (left, red) and human mitoribosomes (right, blue) showing a view of the polypeptide exit site below. The tunnel exit is marked with an asterisk. The polypeptide exit tunnel in mt-LSU is more proteinaceous than in bacteria as a result of two rRNA deletions. (C) Deletion of h7 in bacteria (gray) is compensated by changes to uL29 and an N-terminal extension of uL24. (D) Deletion of h24 (gray) results in the conserved β hairpin of uL24 rotating closer to the tunnel exit and exposes uL22 to the nascent polypeptide.
🎨 Hücre protein fabrikasının özsel bir parçası olarak ribozomal motor
Hücre protein fabrikasının özsel bir parçası olarak ribozomal motor (LINK)
Uzatma etmeni G ya da EF-G'nin bölümleri (yeşil, sarı ve kırmızı) aşağı yukarı motor gibi davranır ve tRNAyı her bir amino asidin eklenmesinin ardından ileriye doğru devinmeye zorlar — gelişmekte olan polipeptid zincire ekleme yapmak için zorunlu bir adım.
Portions of EF-G (in green, yellow and red; elongation factor G) function something like a motor and force tRNA to move forward after the addition of each amino acid, a necessary step to add to the developing polypeptide chain.
📹 What are Ribosomes? — Ribosome Function and Structure / 2min (VİDEO)
📹 What are Ribosomes? — Ribosome Function and Structure / 2min (LINK)
Ribosomes are an underrated hero of the cell and the average mammalian cell has about 10 million ribosomes! They are made of RNA and protein and their function in the cell is to read RNA and synthesize protein, very meta.
Ribosomes have two major subunits, simply called, the large and small ribosomal subunits. Both subunits contain protein and RNA. The RNA in ribosomes is called (not surprisingly) ribosomal RNA, or rRNA for short.
In eukaryotic cells, ribosomes are synthesized in the nucleolus of the nucleus, this is an area densely packed with DNA and proteins. The location of ribosomes in the cell determines where the protein they make ends up. Ribosomes free floating in the cytoplasm of the cell synthesize proteins to be used within the cell, while ribosomes bound to the Rough Endoplasmic Reticulum synthesize proteins that will end up outside the cell, either bound to the cell membrane or released from the cell as a signaling protein for example.
Ribosomes synthesize protein in a process called “translation” where they read strands of mRNA and bring the right amino acids together to build a long polypeptide chain that that will eventually become a functioning protein. You can learn all about transcription and translation here.
📹 mRNA Translation (Advanced) / DNA Learning Center (VİDEO)
📹 mRNA Translation (Advanced) / DNA Learning Center (LINK)
📂 TRANSCRIPT
TRANSCRIPT: The job of this mRNA is to carry the genes message from the DNA out of the nuceus to a ribosome for production of the particular protein that this gene codes for. There can be several million ribosomes in a typical eukaryotic cell these complex catalytic machines use the mrna copy of the genetic information to assemble amino acid building blokes into the three dimensional proteins that are essential for life. Lets see how it works. The ribosome is composed of one large and one small sub-unit that assemble around the messenger RNA, which then passes through the ribosome like a computer tape. The amino acid building blocks (that's the small glowing red molecules) are carried into the ribosome attached to specific transfer RNAs. That's the larger green molecules also referred to as tRNA. The small sub-unit of the ribosome positions the mRNA so that it can be read in groups of three letters known as a codon. Each codon on the mRNA matches a corresponding anti-codon on the base of a transfer RNA molecule.The larger sub-unit of the ribosome removes each amino acid and join it onto the growing protein chain. As the mRNA is ratcheted through the ribosome, the mRNA sequence is translated into an amino acid sequence. There are three locations inside the ribosome, designated the A-site, the P-site and the E-site. The addition of each amino acid is a three step cycle: First, the tRNA enters the ribosome at the A-site and is tested for a codon/anti-codon match with the mRNA. Next, provided there is a correct match, the tRNA is shifted to the P-site and the amino acid it carries is added to the end of the amino acid chain. The mRNA is also ratcheted on three nucleotides or one codon. Thirdly, the spent tRNA is moved to the E-site and then ejected from the ribosome to be recycled. As the protein synthesis proceeds, the finished chain emerges from the ribosome. It folds up into a precise shape, determined by the exact order of amino acids. Thus the Central Dogma explains how the four letter DNA code is - quite literally - turned into flesh and blood.
Within our cells, we refer to ribosomes as the protein factories. Ribosomes are the cellular machines that carry out translation, in this tutorial we will focus on the basics of it's structure and function, as well as the several locations we will find them.
📹 Bacterial ribosome translating RNA into protein / Chemistry World (VİDEO)
📹 Bacterial ribosome translating RNA into protein / Chemistry World (LINK)
This video is from Venki Ramakrishnan's group at the LMB in Cambridge, UK. It shows the ribosome in action, translating RNA into protein. Ramakrishnan won the 2009 Nobel prize for chemistry for his role in working out the structure and function of ribosomes
📹 The Origins and Evolution of the Ribosome / RiboZones (VİDEO)
📹 The Origins and Evolution of the Ribosome / RiboZones (LINK)
The origins and evolution of the ribosome, 3-4 billion years ago, remain imprinted in the biochemistry of extant life and in the structure of the ribosome. This movie takes you back in to time to primordial ribosomes.
📹 Central Dogma & Origin of the Ribosome / Ribozones(VİDEO)
📹 Central Dogma & Origin of the Ribosome / Ribozones (LINK)
This video explains the Central Dogma of Molecular Biology, molecular symbiosis, and the origin of the translation system. The translation system, in analogy with a tree, contains a record of its history. The information contained within ribosomes connects us to the prehistory of biology, spanning 4 billion years of life on earth. Ribosomal RNA variation, observed by comparing three-dimensional structures of ribosomes across the tree of life, forms the basis of our molecular-level model. We infer many steps in the evolution of translation, mapping out acquisition of structure and function, revealing much about how modern biology originated from ancestral chemical systems.
📹 Protein Synthesis Animation Video / Study Force (VİDEO)
📹 Protein Synthesis Animation Video / Study Force (LINK)
📹📹📹 RIBOSOME, PROTEIN SYNTHESIS (VİDEO)
📹 Bacterial Protein synthesis Animation — Initiation, Elongation and Termination / Animated Medical Videos (VİDEO)
📹 Bacterial Protein Synthesis — Initiation, Elongation and Termination / Animated Medical Videos (LINK)
📂 TRANSCRIPTION
Bacterial Protein synthesis Animation - Initiation , Elongation and Termination ( Microbiology )
Gene expression is the process by which genetic information flows from DNA to RNA to the protein. The translation of DNA into RNA is termed transcription; protein synthesis from RNA templates is called translation. Details on gene expression and transcription can be found in a separate learning card.
Translation is carried out by ribosomes, which are large molecular complexes of ribosomal RNA (rRNA) and proteins. Ribosomes bind to RNA templates, also termed messenger RNA (mRNA), and catalyze the formation of a polypeptide based on this template. In the process, a charged transfer RNA (tRNA) recognizes a nucleotide triplet of mRNA that matches a specific amino acid (AA). The new AA is then linked to the next AA of the growing polypeptide on the ribosome. Translation ends once a specific nucleotide sequence of the mRNA is reached (a stop codon). The ribosome subsequently dissociates and the mRNA and newly synthesized protein are released.
Before proteins are functional, a proper shape and destination are both necessary. Proteins begin to fold into their three-dimensional structure during translation according to the AA sequence and local chemical forces and reactions. Various specialized proteins (folding catalysts, chaperones) also help the newly formed proteins to fold properly and reach their correct destinations (e.g., cytosol, organelles, extracellular matrix) via protein modifications. The translation rate of proteins is adjusted to the current conditions of the cell and bodily demands, and is affected by the presence or absence of certain nutrients.
📹 Protein Synthesis (Translation) in Prokaryotes / Alila Medical Media (VİDEO)
📹 Protein Synthesis (Translation) in Prokaryotes / Alila Medical Media (LINK)
📂 TRANSCRIPTION
Initiation of translation in prokaryotes:
The small ribosomal subunit is separated from the large subunit with the help of two initiation factors: IF1 and IF3. This complex then binds a to purine-rich region -- the Shine-Dalgarno sequence -- upstream of the AUG start codon on the mRNA. The Shine-Dalgarno sequence is base-paired to a complementary sequence on the 16S rRNA - a component of the small subunit. This alignment ensures that the start codon is in the right position within the ribosome. Another initiation factor - IF2 - brings in the initiator tRNA charged with the initiator amino acid N-formyl-methionine. The large ribosomal subunit joins the complex and all initiation factors are released. The ribosome has three sites: the A-site is the entry site for new tRNA charged with amino-acid or aminoacyl-tRNA; the P-site is occupied by peptidyl-tRNA - the tRNA that carries the growing polypeptide chain; the E-site is the exit site for the tRNA after it's done delivering the amino acid. The initiator tRNA is positioned in the P-site.
Elongation: A new tRNA carrying an amino acid enters the A-site of the ribosome. On the ribosome, the anticodon of the incoming tRNA is matched against the mRNA codon positioned in the A-site. During this proof-reading, tRNA with incorrect anticodons are rejected and replaced by new tRNA that are again checked. When the right aminoacyl-tRNA enters the A-site, a peptide bond is made between the two now-adjacent amino-acids. As the peptide bond is formed, the tRNA in the P-site releases the amino-acids onto the tRNA in the A-site and becomes empty. At the same time, the ribosome moves one triplet forward on the mRNA. As a result, the empty tRNA is now in the E-site and the peptidyl tRNA is in the P-site. The A-site is now unoccupied and is ready to accept a new tRNA. The cycle is repeated for each codon on the mRNA.
Termination: Termination happens when one of the three stop codons is positioned in the A-site. No tRNA can fit in the A-site at that point as there are no tRNA that match the sequence. Instead, these codons are recognized by a protein, a release factor. Binding of the release factor catalyzes the cleavage of the bond between the polypeptide and the tRNA. The polypeptide is released from the ribosome. The ribosome is disassociated into subunits and is ready for a new round of translation. The newly made polypeptide usually requires additional modifications and folding before it can become an active protein.
📹 Eukaryotic Translation (Protein Synthesis) / Alila Medical Media (VİDEO)
📹 Eukaryotic Translation (Protein Synthesis) / Alila Medical Media (LINK)
📂 TRANSCRIPTION
The translation process involves the following components:
- mRNA or messenger RNA containing the genetic information to be translated.
- tRNA or transfer RNA bringing in the amino acids – the building blocks of the protein.
- Ribosome – the machine that performs the translation. The ribosome has two subunits: small and large.
- Several initiation factors, elongation factors, and release factors. These factors assist with initiation, elongation and termination of the process, respectively.
Steps of the translation process:
Initiation (eukaryotes) : The small ribosomal subunit binds to the initiator tRNA carrying the initiator amino acid methionine. This complex then attaches to the cap structure at the 5’ end of an mRNA and scans for the start codon AUG. The process is mediated by several initiation factors. At the start codon, the large ribosomal subunit joins the complex and all initiation factors are released. The ribosome has three sites: the A-site is the entry site for new tRNA charged with amino-acid or aminoacyl-tRNA; the P-site is occupied by peptidyl-tRNA - the tRNA that carries the growing polypeptide chain; the E-site is the exit site for the tRNA after it’s done delivering the amino acid. The initiator tRNA is positioned in the P-site.
Elongation: A new tRNA carrying an amino acid enters the A-site of the ribosome. On the ribosome, the anticodon of the incoming tRNA is matched against the mRNA codon positioned in the A-site. During this proof-reading, tRNA with incorrect anticodons are rejected and replaced by new tRNA that are again checked. When the right aminoacyl-tRNA enters the A-site, a peptide bond is made between the two now-adjacent amino-acids. As the peptide bond is formed, the tRNA in the P-site releases the amino-acids onto the tRNA in the A-site and becomes empty. At the same time, the ribosome moves one triplet forward on the mRNA. As a result, the empty tRNA is now in the E-site and the peptidyl tRNA is in the P-site. The A-site is now unoccupied and is ready to accept a new tRNA. The cycle is repeated for each codon on the mRNA.
Termination: Termination happens when one of the three stop codons is positioned in the A-site. No tRNA can fit in the A-site at that point as there are no tRNA that match the sequence. Instead, these codons are recognized by a protein, a release factor. Binding of the release factor catalyzes the cleavage of the bond between the polypeptide and the tRNA. The polypeptide is released from the ribosome. The ribosome is disassociated into subunits and is ready for a new round of translation. The newly made polypeptide usually requires additional modifications and folding before it can become an active protein.
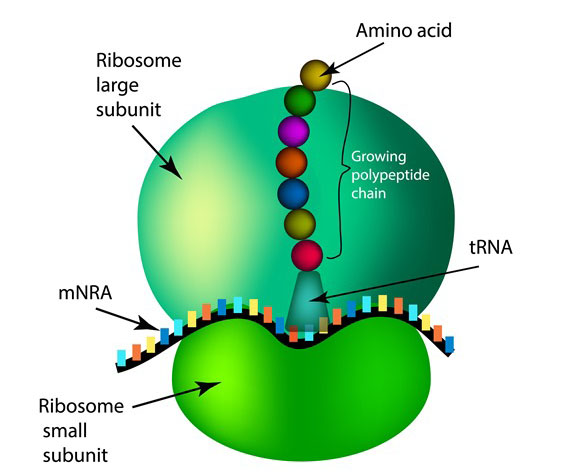
Ribosomes are macromolecular machines, found within all living cells, that perform biological protein synthesis (mRNA translation). Ribosomes link amino acids together in the order specified by the codons of messenger RNA (mRNA) molecules to form polypeptide chains. Ribosomes consist of two major components: the small and large ribosomal subunits. Each subunit consists of one or more ribosomal RNA (rRNA) molecules and many ribosomal proteins (RPs or r-proteins). The ribosomes and associated molecules are also known as the translational apparatus.
Figure 1: Ribosomes assemble polymericproteinmolecules whose sequence is controlled by the sequence of messenger RNA molecules. This is required by all living cells and associated viruses.
The sequence of DNA that encodes the sequence of the amino acids in a protein, is transcribed into a messenger RNA chain. Ribosomes bind to messenger RNAs and use its sequence for determining the correct sequence of amino acids to generate a given protein. Amino acids are selected and carried to the ribosome by transfer RNA (tRNA) molecules, which enter the ribosome and bind to the messenger RNA chain via an anti-codon stem loop. For each coding triplet in the messenger RNA, there is a transfer RNA that matches and carries the correct amino acid for incorporating into a growing polypeptide chain. Once the protein is produced, it can then fold to produce a functional three-dimensional structure.
A ribosome is made from complexes of RNAs and proteins and is therefore a ribonucleoprotein complex. Each ribosome is divided into two subunits:
a smaller subunit which binds to a larger subunit and the mRNA, (30S) and
a larger subunit which binds to the aminoacylated tRNAs and the smaller subunit (50S).
When a ribosome finishes reading an mRNA molecule, these two subunits split apart. Ribosomes are ribozymes,because the catalyticpeptidyl transferase activity that links amino acids together is performed by the ribosomal RNA. Ribosomes are often associated with the intracellular membranes that make up the rough endoplasmic reticulum.
Ribosomes frombacteria,archaea and eukaryotes in the three-domain system resemble each other to a remarkable degree, evidence of a common origin. They differ in their size, sequence, structure, and the ratio of protein to RNA. The differences in structure allow some antibiotics to kill bacteria by inhibiting their ribosomes, while leaving human ribosomes unaffected. In all species, more than one ribosome may move along a single mRNA chain at one time (as a polysome), each "reading" its sequence and producing a corresponding protein molecule.
The mitochondrial ribosomesof eukaryotic cells functionally resemble many features of those in bacteria, reflecting the likely evolutionary origin of mitochondria.
Ribosomes were first observed in the mid-1950s by Romanian-American cell biologist George Emil Palade, using an electron microscope, as dense particles or granules. The term "ribosome" was proposed by scientist Richard B. Roberts in the end of 1950s:
“During the course of the symposium a semantic difficulty became apparent. To some of the participants, "microsomes" mean the ribonucleoprotein particles of the microsome fraction contaminated by other protein and lipid material; to others, the microsomes consist of protein and lipid contaminated by particles. The phrase "microsomal particles" does not seem adequate, and "ribonucleoprotein particles of the microsome fraction" is much too awkward. During the meeting, the word "ribosome" was suggested, which has a very satisfactory name and a pleasant sound. The present confusion would be eliminated if "ribosome" were adopted to designate ribonucleoprotein particles in sizes ranging from 35 to 100S.”
— Albert, Microsomal Particles and Protein Synthesis
Figure 2: Large (red) and small (blue) subunit fit together.
The ribosome is a highly complex cellular machine. It is largely made up of specialized RNA known as ribosomal RNA (rRNA) as well as dozens of distinct proteins (the exact number varies slightly between species). The ribosomal proteins and rRNAs are arranged into two distinct ribosomal pieces of different size, known generally as the large and small subunit of the ribosome. Ribosomes consist of two subunits that fit together (Figure 2) and work as one to translate the mRNA into a polypeptide chain during protein synthesis (Figure 1). Because they are formed from two subunits of non-equal size, they are slightly longer in the axis than in diameter.
Prokaryotic ribosomes are around 20 nm (200 Å) in diameter and are composed of 65% rRNA and 35% ribosomal proteins. Eukaryotic ribosomes are between 25 and 30 nm (250–300 Å) in diameter with an rRNA-to-protein ratio that is close to 1. Crystallographic work has shown that there are no ribosomal proteins close to the reaction site for polypeptide synthesis. This suggests that the protein components of ribosomes do not directly participate in peptide bond formation catalysis, but rather that these proteins act as a scaffold that may enhance the ability of rRNA to synthesize protein (See: Ribozyme).
The ribosomal subunits of bacteria and eukaryotes are quite similar.
The unit of measurement used to describe the ribosomal subunits and the rRNA fragments is the Svedberg unit, a measure of the rate of sedimentation in centrifugation rather than size. This accounts for why fragment names do not add up: for example, bacterial 70S ribosomes are made of 50S and 30S subunits.
Bacteria have 70S ribosomes, each consisting of a small (30S) and a large (50S) subunit. E. coli, for example, has a 16S RNA subunit (consisting of 1540 nucleotides) that is bound to 21 proteins. The large subunit is composed of a 5S RNA subunit (120 nucleotides), a 23S RNA subunit (2900 nucleotides) and 31 proteins.
Affinity label for the tRNA binding sites on the E. coli ribosome allowed the identification of A and P site proteins most likely associated with the peptidyltransferase activity; labelled proteins are L27, L14, L15, L16, L2; at least L27 is located at the donor site, as shown by E. Collatz and A.P. Czernilofsky. Additional research has demonstrated that the S1 and S21 proteins, in association with the 3′-end of 16S ribosomal RNA, are involved in the initiation of translation.
Figure 3: Atomic structure of the 30S subunit from Thermus thermophilus. Proteins are shown in blue and the single RNA chain in brown.
Eukaryotes have 80S ribosomes located in their cytosol, each consisting of a small (40S) and large (60S) subunit. Their 40S subunit has an 18S RNA (1900 nucleotides) and 33 proteins. The large subunit is composed of a 5S RNA (120 nucleotides), 28S RNA (4700 nucleotides), a 5.8S RNA (160 nucleotides) subunits and 46 proteins.
eukaryotic cytosolic ribosomes (R. norvegicus)
ribosome
subunit
rRNAs
r-proteins
80S
60S
28S (4718 nt)
49
5.8S (160 nt)
5S (120 nt)
40S
18S (1874 nt)
33
During 1977, Czernilofsky published research that used affinity labeling to identify tRNA-binding sites on rat liver ribosomes. Several proteins, including L32/33, L36, L21, L23, L28/29 and L13 were implicated as being at or near the peptidyl transferase center.
In eukaryotes, ribosomes are present in mitochondria (sometimes called mitoribosomes) and in plastids such as chloroplasts (also called plastoribosomes). They also consist of large and small subunits bound together with proteins into one 70S particle. These ribosomes are similar to those of bacteria and these organelles are thought to have originated as symbioticbacteria Of the two, chloroplastic ribosomes are closer to bacterial ones than mitochrondrial ones are. Many pieces of ribosomal RNA in the mitochrondria are shortened, and in the case of 5S rRNA, replaced by other structures in animals and fungi. In particular, Leishmania tarentolae has a minimalized set of mitochondrial rRNA.
The cryptomonad and chlorarachniophyte algae may contain a nucleomorph that resembles a vestigial eukaryotic nucleus. Eukaryotic 80S ribosomes may be present in the compartment containing the nucleomorph.
The differences between the bacterial and eukaryotic ribosomes are exploited by pharmaceutical chemists to create antibiotics that can destroy a bacterial infection without harming the cells of the infected person. Due to the differences in their structures, the bacterial 70S ribosomes are vulnerable to these antibiotics while the eukaryotic 80S ribosomes are not. Even though mitochondria possess ribosomes similar to the bacterial ones, mitochondria are not affected by these antibiotics because they are surrounded by a double membrane that does not easily admit these antibiotics into the organelle. A noteworthy counterexample, however, includes the antineoplastic antibiotic chloramphenicol, which successfully inhibits bacterial 50S and mitochondrial 50S ribosomes. The same of mitochondria cannot be said of chloroplasts, where antibiotic resistance in ribosomal proteins is a trait to be introduced as a marker in genetic engineering
The various ribosomes share a core structure, which is quite similar despite the large differences in size. Much of the RNA is highly organized into various tertiary structural motifs, for example pseudoknots that exhibit coaxial stacking. The extra RNA in the larger ribosomes is in several long continuous insertions, such that they form loops out of the core structure without disrupting or changing it. All of the catalytic activity of the ribosome is carried out by the RNA; the proteins reside on the surface and seem to stabilize the structure.
The general molecular structure of the ribosome has been known since the early 1970s. In the early 2000s, the structure has been achieved at high resolutions, of the order of a few ångströms.
The first papers giving the structure of the ribosome at atomic resolution were published almost simultaneously in late 2000. The 50S (large prokaryotic) subunit was determined from the archaeonHaloarcula marismortui and the bacterium Deinococcus radiodurans, and the structure of the 30S subunit was determined from Thermus thermophilus. These structural studies were awarded the Nobel Prize in Chemistry in 2009. In May 2001 these coordinates were used to reconstruct the entire T. thermophilus 70S particle at 5.5 Å resolution.
Two papers were published in November 2005 with structures of the Escherichia coli 70S ribosome. The structures of a vacant ribosome were determined at 3.5 Å resolution using X-ray crystallography. Then, two weeks later, a structure based on cryo-electron microscopy was published, which depicts the ribosome at 11–15 Å resolution in the act of passing a newly synthesized protein strand into the protein-conducting channel.
The first atomic structures of the ribosome complexed with tRNA and mRNA molecules were solved by using X-ray crystallography by two groups independently, at 2.8 Å and at 3.7 Å. These structures allow one to see the details of interactions of the Thermus thermophilus ribosome with mRNA and with tRNAs bound at classical ribosomal sites. Interactions of the ribosome with long mRNAs containing Shine-Dalgarno sequences were visualized soon after that at 4.5–5.5 Å resolution.
In 2011, the first complete atomic structure of the eukaryotic 80S ribosome from the yeast Saccharomyces cerevisiae was obtained by crystallography. The model reveals the architecture of eukaryote-specific elements and their interaction with the universally conserved core. At the same time, the complete model of a eukaryotic 40S ribosomal structure in Tetrahymena thermophila was published and described the structure of the 40S subunit, as well as much about the 40S subunit's interaction with eIF1 during translation initiation. Similarly, the eukaryotic 60S subunit structure was also determined from Tetrahymena thermophila in complex with eIF6.
Figure 4: Atomic structure of the 50S subunit from Haloarcula marismortui. Proteins are shown in blue and the two RNA chains in brown and yellow. The small patch of green in the center of the subunit is the active site..
Ribosomes are minute particles consisting of RNA and associated proteins that function to synthesize proteins. Proteins are needed for many cellular functions such as repairing damage or directing chemical processes. Ribosomes can be found floating within the cytoplasm or attached to the endoplasmic reticulum. Basically, their main function is to convert genetic code into an amino acid sequence and to build protein polymers from amino acid monomers.
Ribosomes act as catalysts in two extremely important biological processes called peptidyl transfer and peptidyl hydrolysis. The "PT center is responsible for producing protein bonds during protein elongation."
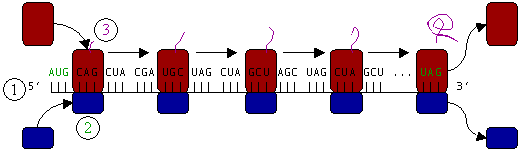
Ribosomes are the workplaces of protein biosynthesis, the process of translating mRNA into protein. The mRNA comprises a series of codons which are decoded by the ribosome so as to make the protein. Using the mRNA as a template, the ribosome traverses each codon (3 nucleotides) of the mRNA, pairing it with the appropriate amino acid provided by an aminoacyl-tRNA. Aminoacyl-tRNA contains a complementary anticodon on one end and the appropriate amino acid on the other. For fast and accurate recognition of the appropriate tRNA, the ribosome utilizes large conformational changes (conformational proofreading) . The small ribosomal subunit, typically bound to an aminoacyl-tRNA containing the first amino acid methionine, binds to an AUG codon on the mRNA and recruits the large ribosomal subunit. The ribosome contains three RNA binding sites, designated A, P and E. The A-site binds an aminoacyl-tRNA or termination release factors; the P-site binds a peptidyl-tRNA (a tRNA bound to the poly-peptide chain); and the E-site (exit) binds a free tRNA. Protein synthesis begins at a start codon AUG near the 5' end of the mRNA. mRNA binds to the P site of the ribosome first. The ribosome recognizes the start codon by using the Shine-Dalgarno sequence of the mRNA in prokaryotes and Kozak box in eukaryotes.
Although catalysis of the peptide bond involves the C2 hydroxyl of RNA's P-site adenosine in a proton shuttle mechanism, other steps in protein synthesis (such as translocation) are caused by changes in protein conformations. Since their catalytic core is made of RNA, ribosomes are classified as "ribozymes," and it is thought that they might be remnants of the RNA world.
In Figure 5, both ribosomal subunits (small and large) assemble at the start codon (towards the 5' end of the mRNA). The ribosome uses tRNA that matches the current codon (triplet) on the mRNA to append an amino acid to the polypeptide chain. This is done for each triplet on the mRNA, while the ribosome moves towards the 3' end of the mRNA. Usually in bacterial cells, several ribosomes are working parallel on a single mRNA, forming what is called a polyribosome or polysome.
Figure 5: Translation of mRNA (1) by a ribosome (2) (shown as small and large subunits) into a polypeptide chain (3). The ribosome begins at the start codon of RNA (AUG) and ends at the stop codon (UAG).
The ribosome is known to actively participate in the protein folding. The structures obtained in this way are usually identical to the ones obtained during protein chemical refolding, however, the pathways leading to the final product may be different. In some cases, the ribosome is crucial in obtaining the functional protein form. For example, one of the possible mechanisms of folding of the deeply knotted proteins relies on the ribosome pushing the chain through the attached loop.
Addition of translation-independent amino acids
Addition of translation-independent amino acids (W)
Presence of a ribosome quality control protein Rqc2 is associated with mRNA-independent protein elongation. This elongation is a result of ribosomal addition (via tRNAs brought by Rqc2) of CAT tails: ribosomes extend the C-terminus of a stalled protein with random, translation-independent sequences of alanines and threonines.
Ribosomes are classified as being either "free" or "membrane-bound".
Free and membrane-bound ribosomes differ only in their spatial distribution; they are identical in structure. Whether the ribosome exists in a free or membrane-bound state depends on the presence of an ER-targeting signal sequence on the protein being synthesized, so an individual ribosome might be membrane-bound when it is making one protein, but free in the cytosol when it makes another protein.
Ribosomes are sometimes referred to as organelles, but the use of the term organelle is often restricted to describing sub-cellular components that include a phospholipid membrane, which ribosomes, being entirely particulate, do not. For this reason, ribosomes may sometimes be described as "non-membranous organelles".
Figure 6: A ribosome translating a protein that is secreted into the endoplasmic reticulum.
Free ribosomes can move about anywhere in the cytosol, but are excluded from the cell nucleus and other organelles. Proteins that are formed from free ribosomes are released into the cytosol and used within the cell. Since the cytosol contains high concentrations of glutathione and is, therefore, a reducing environment, proteins containing disulfide bonds, which are formed from oxidized cysteine residues, cannot be produced within it.
When a ribosome begins to synthesize proteins that are needed in some organelles, the ribosome making this protein can become "membrane-bound". In eukaryotic cells this happens in a region of the endoplasmic reticulum (ER) called the "rough ER". The newly produced polypeptide chains are inserted directly into the ER by the ribosome undertaking vectorial synthesis and are then transported to their destinations, through the secretory pathway. Bound ribosomes usually produce proteins that are used within the plasma membrane or are expelled from the cell via exocytosis.
In bacterial cells, ribosomes are synthesized in the cytoplasm through the transcription of multiple ribosome gene operons. In eukaryotes, the process takes place both in the cell cytoplasm and in the nucleolus, which is a region within the cell nucleus. The assembly process involves the coordinated function of over 200 proteins in the synthesis and processing of the four rRNAs, as well as assembly of those rRNAs with the ribosomal proteins.
The ribosome may have first originated in an RNA world,appearing as a self-replicating complex that only later evolved the ability to synthesize proteins when amino acids began to appear. Studies suggest that ancient ribosomes constructed solely of rRNA could have developed the ability to synthesize peptide bonds. In addition, evidence strongly points to ancient ribosomes as self-replicating complexes, where the rRNA in the ribosomes had informational, structural, and catalytic purposes because it could have coded for tRNAs and proteins needed for ribosomal self-replication. Hypothetical cellular organisms with self-replicating RNA but without DNA are called ribocytes (or ribocells).
As amino acids gradually appeared in the RNA world under prebiotic conditions, their interactions with catalytic RNA would increase both the range and efficiency of function of catalytic RNA molecules. Thus, the driving force for the evolution of the ribosome from an ancient self-replicating machine into its current form as a translational machine may have been the selective pressure to incorporate proteins into the ribosome's self-replicating mechanisms, so as to increase its capacity for self-replication.
Ribosomes are compositionally heterogeneous between species and even within the same cell, as evidenced by the existence of cytoplasmic and mitochondria ribosomes within the same eukaryotic cells. Certain researchers have suggested that heterogeneity in the composition of ribosomal proteins is mammals is important for gene regulation, i.e., the specialized ribosome hypothesis. However, this hypothesis is controversial and the topic of ongoing research.
Heterogeneity in ribosome composition was first proposed to be involved in translational control of protein synthesis by Vince Mauro and Gerald Edelman. They proposed the ribosome filter hypothesis to explain the regulatory functions of ribosomes. Evidence has suggested that specialized ribosomes specific to different cell populations may affect how genes are translated. Some ribosomal proteins exchange from the assembled complex with cytosolic copies suggesting that the structure of the in vivo ribosome can be modified without synthesizing an entire new ribosome.
Certain ribosomal proteins are absolutely critical for cellular life while others are not. In budding yeast, 14/78 ribosomal proteins are non-essential for growth, while in humans this depends on the cell of study. Other forms of heterogeneity include post-translational modifications to ribosomal proteins such as acetylation, methylation, and phosphorylation. Arabidopsis, Viral internal ribosome entry sites (IRESs) may mediate translations by compositionally distinct ribosomes. For example, 40S ribosomal units without eS25 in yeast and mammalian cells are unable to recruit the CrPV IGR IRES.
Heterogeneity of ribosomal RNA modifications plays an important role in structural maintenance and/or function and most mRNA modifications are found in highly conserved regions. The most common rRNA modifications are pseudouridylation and 2’-O methylation of ribose.
Ribosome, particle that is present in large numbers in all living cells and serves as the site of protein synthesis. Ribosomes occur both as free particles in prokaryotic and eukaryotic cells and as particles attached to the membranes of the endoplasmic reticulum in eukaryotic cells. The small particles that came to be known as ribosomes were first described in 1955 by Romanian-born American cell biologist George E. Palade, who found them to be frequently associated with the endoplasmic reticulum in eukaryotic cells.
Ribosomes on the outer surface of the endoplasmic reticulum play an important role in protein synthesis within cells.
Ribosomes are remarkably abundant in cells. A single actively replicating eukaryotic cell, for example, may contain as many as 10 million ribosomes. In the bacteriumEscherichia coli (a prokaryote), ribosomes may number as many as 15,000, constituting as much as one-quarter of the cell’s total mass. The size of the ribosomes within cells varies, depending on the cell type and on factors such as whether the cell is resting or replicating. The average ribosome of E. coli, the best-characterized example, measures about 200 angstroms (about 20 nm) in diameter.
Protein synthesis
DNA in the cell nucleus carries a genetic code, which consists of sequences of adenine (A), thymine (T), guanine (G), and cytosine (C) (Figure 1). RNA, which contains uracil (U) instead of thymine, carries the code to protein-making sites in the cell. To make RNA, DNA pairs its bases with those of the "free" nucleotides (Figure 2). Messenger RNA (mRNA) then travels to the ribosomes in the cell cytoplasm, where protein synthesis occurs (Figure 3). The base triplets of transfer RNA (tRNA) pair with those of mRNA and at the same time deposit their amino acids on the growing protein chain. Finally, the synthesized protein is released to perform its task in the cell or elsewhere in the body.
.
Ribosomes are made up of ribosomal proteins and ribosomal RNA (rRNA). In prokaryotes, ribosomes are roughly 40 percent protein and 60 percent rRNA. In eukaryotes, ribosomes are about half protein and half rRNA. Ribosomes are usually made up of three or four rRNA molecules and anywhere from about 40 to 80 different ribosomal proteins.
Each ribosome is composed of two subunits, a larger one and a smaller one, each of which has a characteristic shape. The subunits typically are referred to in terms of their sedimentation rate, which is measured in Svedberg units (S), in a centrifugal field. The small and large subunits of eukaryotes are designated 40S and 60S, respectively, while prokaryotes contain a small 30S subunit and a large 50S subunit.
Ribosomes are the sites at which information carried in the genetic code is converted into protein molecules. Ribosomal molecules of messenger RNA (mRNA) determine the order of transfer RNA (tRNA) molecules that are bound to nucleotide triplets (codons). The order of tRNA molecules ultimately determines the amino acid sequence of a protein. Molecules of rRNA catalyze the peptidyl transferase reaction, which forms peptide bonds between the amino acids, linking them together to form proteins. The newly formed proteins detach themselves from the ribosome site and migrate to other parts of the cell for use.


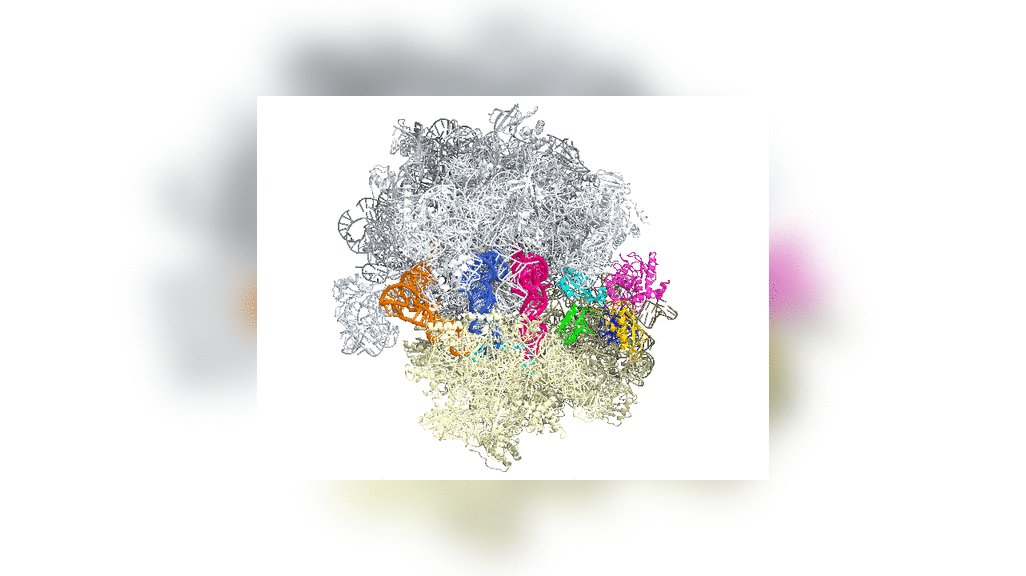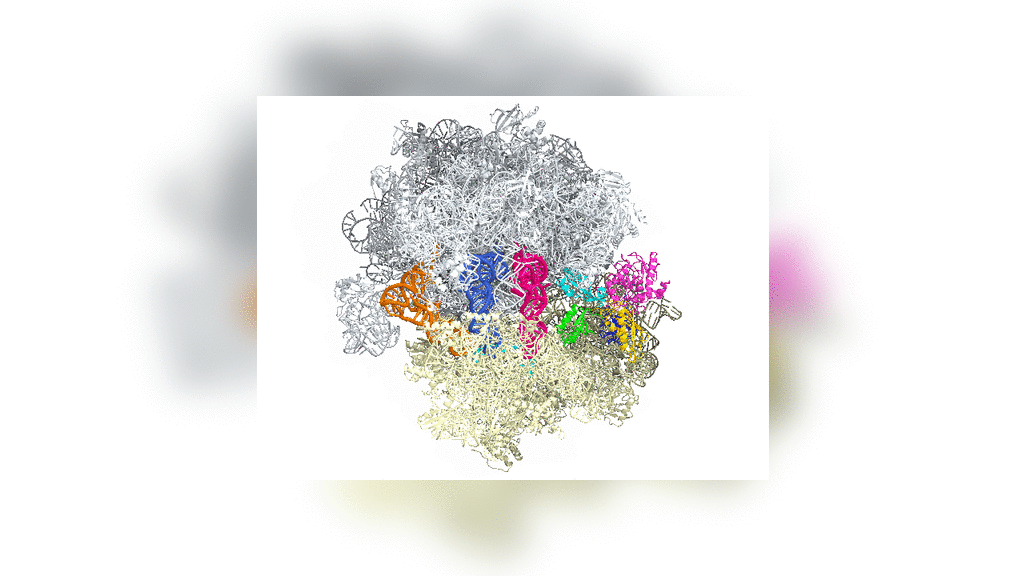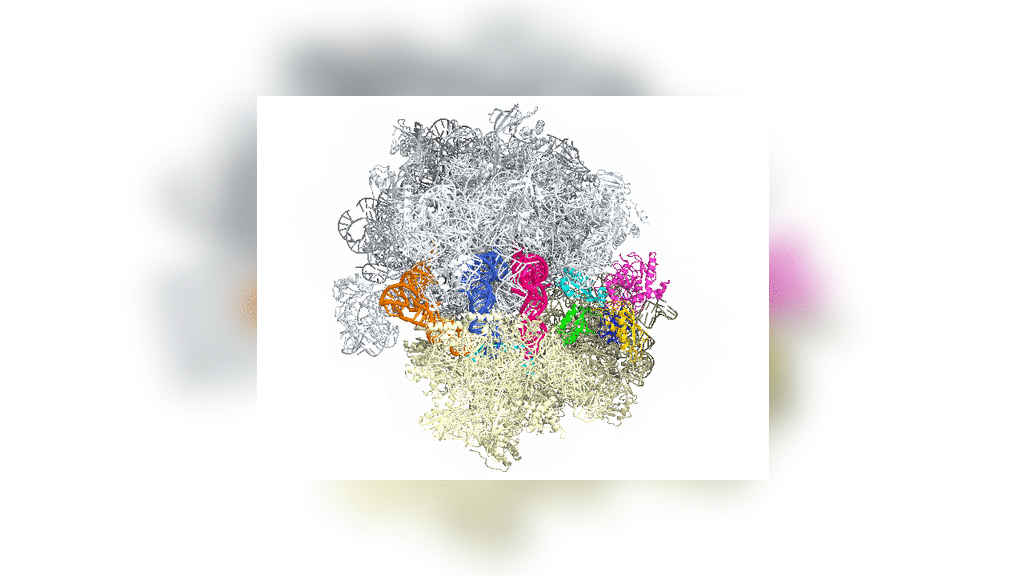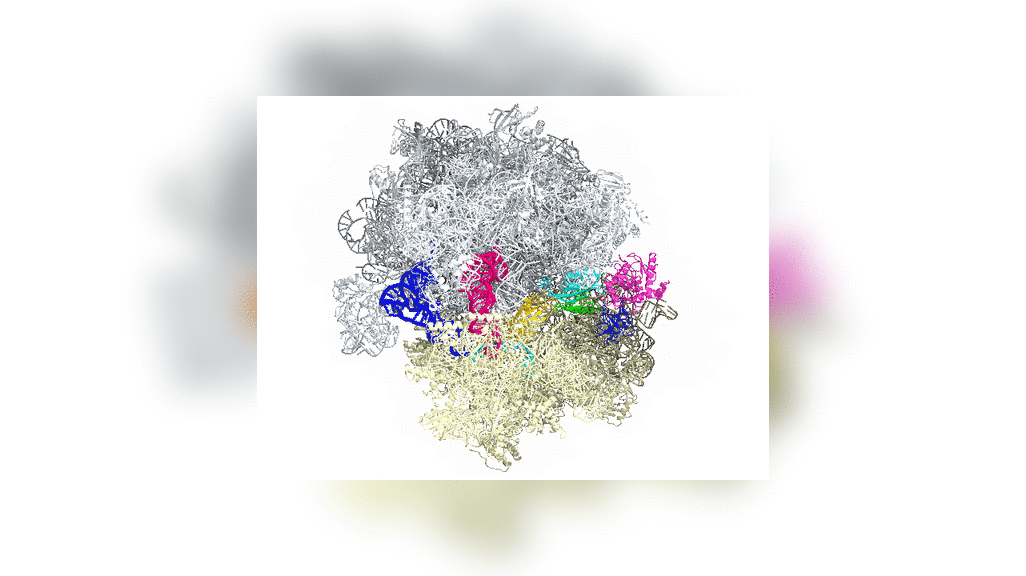


.jpg)