Life, living matter and, as such, matter that shows certain attributes that include responsiveness, growth, metabolism, energy transformation, and reproduction.Although a noun, as with other defined entities, the word life might be better cast as a verb to reflect its essential status as a process.{?} Life comprises individuals, {?} living beings, assignable to groups (taxa). Each individual is composed of one or more minimal living units, called cells, and is capable of transformation of carbon-based and other compounds (metabolism), growth, and participation in reproductive acts. Life-forms present on Earth today have evolved from ancient common ancestors through the generation of hereditary variation and natural selection. Although some studies state that life may have begun as early as 4.1 billion years ago, it can be traced to fossils dated to 3.5-3.7 billion years ago, which is still only slightly younger than Earth, which gravitationally accreted into a planet about 4.5 billion years ago. But this is life as a whole. More than 99.9 percent of species that have ever lived are extinct. The several branches of science that reveal the common historical, functional, and chemical basis of the evolution of all life include electron microscopy,genetics, paleobiology (including paleontology), and molecular biology.
African elephants (Loxodonta africana) in Botswana.
The phenomenon of life can be approached in several ways: life as it is known and studied on planet Earth; life imaginable in principle; and life, by hypothesis, that might exist elsewhere in the universe (seeextraterrestrial life). As far as is known, life exists only on Earth. Most life-forms reside in a thin sphere that extends about 23 km (14 miles) from 3 km (2 miles) beneath the bottom of the ocean to the top of the troposphere (lower atmosphere); the relative thickness is comparable to a coat of paint on a rubber ball. An estimated 10–30 million distinguishable species currently inhabit this sphere of life, or biosphere.
Mangroves (Rhizophora apiculata) at low tide on the coast of Thailand.
Definitions Of Life
Definitions Of Life
Definitions Of Life (B)
Much is known about life from points of view reflected in the various biological, or “life,” sciences. These include anatomy (the study of form at the visible level), ultrastructure (the study of form at the microscopic level), physiology (the study of function), molecular biology and biochemistry (the study of form and function at chemical levels), ecology (the study of the relations of organisms with their environments), taxonomy (the naming, identifying, and classifying of organisms), ethology (the study of animal behaviour), and sociobiology (the study of social behaviour). Specific sciences that participate in the study of life focus more narrowly on certain taxa or levels of observation — e.g., botany (the study of plants), lichenology (the study of lichens, leafy or crusty individuals composed of permanent associations between algae or photosyntheticbacteria and fungi),herpetology (the study of amphibians and reptiles),microbiology (the study of bacteria, yeast, and other unicellular fungi, archaea, protists,viruses),zoology (the study of marine and land animals), and cytology (the study of cells). Although the scientists, technicians, and others who participate in studies of life easily distinguish living matter from inert or dead matter, none can give a completely inclusive, concise definition of life itself. Part of the problem is that the core properties of life — growth, change, reproduction, active resistance to external perturbation, and evolution — involve transformation or the capacity for transformation. Living processes are thus antithetical to a desire for tidy classification or final definition. To take one example, the number of chemical elements involved with life has increased with time; an exhaustive list of the material constituents of life would therefore be premature. Nonetheless, most scientists implicitly use one or more of the metabolic, physiological, biochemical, genetic, thermodynamic, and autopoietic definitions given below.
Metabolic
Metabolic (B) Metabolic definitions are popular with biochemists and some biologists. Living systems are objects with definite boundaries, continually exchanging some materials with their surroundings but without altering their general properties, at least over some period of time. However, there are exceptions. There are frozen seeds and spores that remain, so far as is known, perfectly dormant. At low temperatures they lack metabolic activity for hundreds, perhaps thousands, of years but revive perfectly well upon being subjected to more clement conditions. A candle flame has a well-defined shape with a fixed boundary and is maintained by “metabolizing” its organic waxes and the surrounding molecular oxygen to produce carbon dioxide and water. Similar reactions, incidentally, occur in animals and plants. Flames also have a well-known capacity for growth. These facts underscore the inadequacy of this metabolic definition, even as they suggest the indispensable role of energy transformation to living systems. (Seemetabolism.)
Physiological
Physiological (B) Physiological definitions of life are popular. Life is defined as any system capable of performing functions such as eating, metabolizing, excreting, breathing, moving, growing, reproducing, and responding to external stimuli. But many such properties are either present in machines that nobody is willing to call alive or absent from organisms, such as the dormant hard-covered seed of a tree, that everybody is willing to call alive. An automobile, for example, can be said to eat, metabolize, excrete, breathe, move, and be responsive to external stimuli. A visitor from another planet, judging from the enormous number of automobiles on Earth and the way in which cities and landscapes have been designed for the special benefit of motorcars, might well believe that automobiles are not only alive but are the dominant life-form on the planet. (Seephysiology.)
Active traps of the Venus's-flytrap (Dionaea muscipula), a carnivorous plant. If depressed at least twice, thin pressure-sensitive hairs in the trap stimulate the lobes to clamp tightly over an insect.
Biochemical
Biochemical (B)
A biochemical or molecular biological definition sees living organisms as systems that contain reproducible hereditary information coded in nucleic acidmolecules and that metabolize by controlling the rate of chemical reactions using the proteinaceous catalysts known as enzymes. In many respects, this is more satisfying than the physiological or metabolic definitions of life. However, even here there are counterexamples. Viruslike agents called prions lack nucleic acids, although the nucleic acids of the animal cells in which they reside may be involved in their reproduction. Ribonucleic acid (RNA) molecules may replicate, mutate, and then replicate their mutations in test tubes, although by themselves they are not alive. Furthermore, a definition strictly in chemical terms seems peculiarly vulnerable. It implies that, were a person able to construct a system that had all the functional properties of life, it would still not be alive if it lacked the molecules that earthly biologists are fond of—and made of. (Seebiochemistry.)
Genetic
Genetic (B)
All organisms on Earth, from the tiniest cell to the loftiest trees, display extraordinary powers. They effortlessly perform complex transformations of organic molecules, exhibit elaborate behaviour patterns, and indefinitely construct from raw materials in the environment more or less identical copies of themselves. How could systems of such staggering complexity and such stunning beauty ever arise? A main part of the answer, for which today there is excellent scientific evidence, was carefully chronicled by the English naturalist Charles Darwin in the years before the publication in 1859 of his epoch-making work On the Origin of Species. A modern rephrasing of his theory of natural selection goes something like this: Hereditary information is carried by large molecules known as genes, composed of nucleic acids. Different genes are responsible for the expression of different characteristics of the organism. During the reproduction of the organism, the genes also replicate and thereby pass on the instructions for various characteristics to the next generation. Occasionally, there are imperfections, called mutations, in gene replication. A mutation alters the instructions for one or more particular characteristics. The mutation also breeds true, in the sense that its capability for determining a given characteristic of the organism remains unimpaired for generations until the mutated gene is itself mutated. Some mutations, when expressed, will produce characteristics favourable for the organism; organisms with such favourable genes will reproduce preferentially over those without such genes. Most mutations, however, turn out to be deleterious and often lead to some impairment or to death of the organism. (To illustrate, it is unlikely that one can improve the functioning of a finely crafted watch by dropping it from a tall building. The watch may perform better, but this is highly improbable.) In this way, organisms slowly evolve toward greater complexity. This evolution occurs, however, only at enormous cost: modern humans, complex and reasonably well-adapted, exist only because of billions of deaths of organisms slightly less adapted and somewhat less complex. In short, Darwin’s theory of natural selection states that complex organisms evolved through time because of replication, mutation, and replication of mutations. A genetic definition of life therefore would be a system capable of evolution by natural selection. (SeeDarwinism.)
The initial proposal of the structure of DNA by James Watson and Francis Crick, which was accompanied by a suggestion on the means of replication.
This definition places great emphasis on the importance of replication.Replication refers to the capacity of molecules such as deoxyribonucleic acid (DNA) to precisely copy themselves, whereas reproduction refers to the increase in number of organisms by acts that make a new individual from its parent or parents. In any organism, enormous biological effort is directed toward reproduction, although it confers no obvious benefit on the reproducing organism itself. However, if life is defined as an entity capable of reproduction, then a mule, which is clearly alive yet does not reproduce, would be excluded from the living under this restrictive definition. Indeed, many organisms, such as hybrid mammals and plants that are past their prime, do not reproduce even though the individual cells of which they are composed may.
Life defined as a reproductive system dependent on replicating components does not rule out synthetic reproduction. For example, it should be possible to construct a machine that is capable of producing identical copies of itself from preformed building blocks but that arranges its descendants in a slightly different manner when a random change occurs in its instructions. Such a machine would of course reproduce its instructions as well. But the fact that such a machine would satisfy the genetic definition of life is not an argument against such a definition; in fact, if the building blocks were simple enough, the machine would have the capability of evolving into very complex systems that would probably have all the other properties attributed to living systems. (Some computer programmers have already claimed, on the basis of running generations of replicating and mutating computer instructions, to have created artificial life [“a-life”]; such programs do not, however, show any real freedom or awareness, and their activities are thus far limited to the insides of computers.) The genetic definition has the additional advantage of being expressed purely in functional terms; i.e., it does not depend on any particular choice of constituent molecules. The improbability of contemporary organisms—dealt with more fully below—is so great that these organisms could not possibly have arisen by purely random processes and without historical continuity. Fundamental to the genetic definition of life then seems to be the notion that a certain level of complexity cannot be achieved without natural selection.
Thermodynamic
Thermodynamic (B) Thermodynamics distinguishes between isolated, closed, and open systems. An isolated system is separated from the rest of the environment and exchanges neither light nor heat nor matter with its surroundings. A closed system exchanges energy but not matter. An open system is one in which both material and energetic exchanges occur. The second law of thermodynamics states that, in a closed system, no processes will tend to occur that increase the net organization (or decrease the net entropy) of the system. Thus, the universe taken as a whole is steadily moving toward a state of complete randomness, lacking any order, pattern, or beauty. This fate was popularized in the 19th century as the “heat death” of the universe.
Living organisms are manifestly organized and at first sight seem to represent a contradiction to the second law of thermodynamics. Indeed, living systems might then be defined as localized regions where there is a continuous maintenance or increase in organization. Living systems, however, do not really contradict the second law. They increase their organization in regions of energy flow, and, indeed, their cycling of materials and their tendency to grow can be understood only in the context of a more general definition of the second law that applies to open as well as closed and isolated systems. In nature (except at cosmic scales, where gravity becomes a crucial factor), energy moves from being concentrated to being spread out; spontaneously occurring complex systems do not violate the second law but help energy spread out, thus producing entropy and reducing gradients.
A general statement of open-system thermodynamics is that nature abhors a gradient, a difference across a distance. Differences and gradients in nature represent improbable, preexisting organizations. Many complex systems in nature spontaneously arise to degrade gradients and persist until the gradients are nullified. A tornado, for example, is an improbable, matter-cycling system that appears in the area of a barometric pressure gradient; when the air pressure gradient is gone, the “improbable” tornado disappears. Life seems to be a similar system, but one that degrades the solar gradient, the electromagnetic difference between the extremely hot (5,800 K [5,500 °C, or 10,000 °F]) Sun and very cold (2.7 K [−270.3 °C, or −454.5 °F]) outer space. (K = kelvin. On the Kelvin temperature scale, in which 0 K [−273 °C, or −460 °F] is absolute zero, 273 K [0 °C, or 32 °F] is the freezing point of water, and 373 K [100 °C, or 212 °F] is the boiling point of water at one atmosphere pressure.) Most life on Earth is dependent on the flow of sunlight, which is utilized by photosynthetic organisms to construct complex molecules from simpler ones. Some deep-sea and cave organisms called chemoautotrophs depend on chemical gradients, such as the natural energy-producing reaction between hydrogen sulfide bubbling up from vents and oxygen dissolved in water. The organization of life on Earth can thus be seen as being driven by a natural second-law-based reduction between the energy of the hot Sun and the cooler space around it. Although life has not fully reduced the solar gradient, incorporation of carbon dioxide into chemoautotrophs and production of clouds by plants help keep Earth’s surface cooler than it would otherwise be, thereby helping to degrade the solar energy gradient.
Some scientists argue on grounds of quite general open-system thermodynamics that the organization of a system increases as energy flows through it. Moreover, energy flow leads to the development of cycles. An example of a biological cycle on Earth is the carbon cycle.Carbon from atmospheric carbon dioxide is incorporated by photosynthetic or chemosynthetic organisms and converted into carbohydrates through the process of autotrophy. These carbohydrates are ultimately oxidized by heterotrophic organisms to extract useful energy locked in their chemical bonds. In the oxidation of carbohydrates, carbon dioxide is returned to the atmosphere, thus completing the cycle. It has been shown that similar cycles develop spontaneously and in the absence of life by the flow of energy through chemical systems. Biological cycles may represent natural thermodynamic cycles reinforced by a genetic apparatus. It seems doubtful that open-system thermodynamic processes in the absence of replication lead to the sorts of complexity that characterize biological systems; replication, however, may be interpreted as an especially efficient thermodynamic means of gradient breakdown—a kind of special, slow-burning “fire.” In any case, it is clear that much of the complexity of life on Earth has arisen through replication, with thermodynamically favoured pathways being used by energy-transforming organisms.
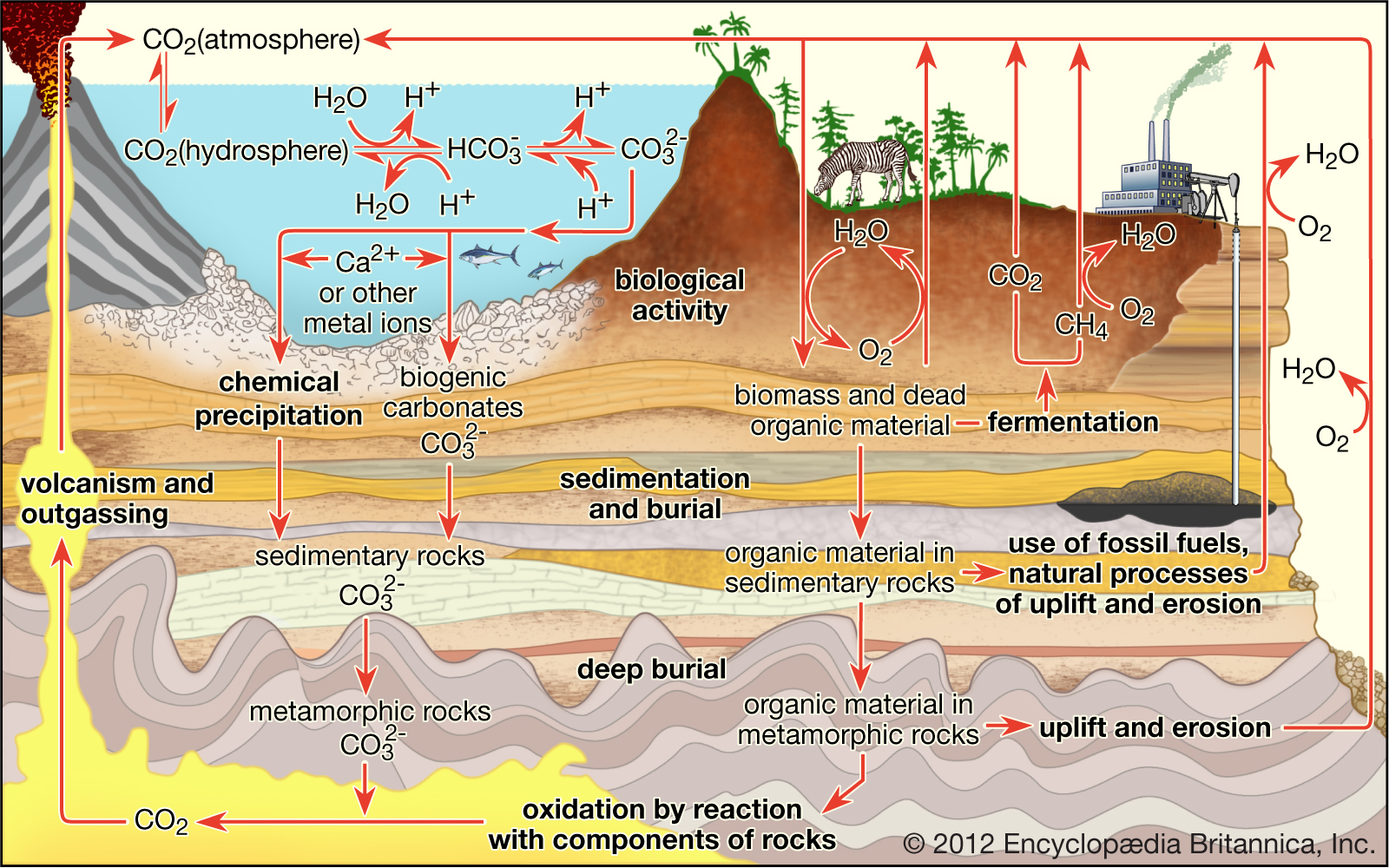
Carbon cycle
Carbon is transported in various forms through the atmosphere, the hydrosphere, and geologic formations. One of the primary pathways for the exchange of carbon dioxide (CO2) takes place between the atmosphere and the oceans; there a fraction of the CO2 combines with water, forming carbonic acid (H2CO3) that subsequently loses hydrogen ions (H+) to form bicarbonate (HCO3−) and carbonate (CO32−) ions. Mollusk shells or mineral precipitates that form by the reaction of calcium or other metal ions with carbonate may become buried in geologic strata and eventually release CO2 through volcanic outgassing. Carbon dioxide also exchanges through photosynthesis in plants and through respiration in animals. Dead and decaying organic matter may ferment and release CO2 or methane (CH4) or may be incorporated into sedimentary rock, where it is converted to fossil fuels. Burning of hydrocarbon fuels returns CO2 and water (H2O) to the atmosphere. The biological and anthropogenic pathways are much faster than the geochemical pathways and, consequently, have a greater impact on the composition and temperature of the atmosphere.
Autopoietic
Autopoietic (B)
A newer definition of life revolves around the idea of autopoiesis. This idea was put forth by Chilean biologists Humberto Maturana and Francisco Varela and emphasizes the peculiar closure of living systems, which are alive and maintain themselves metabolically whether they succeed in reproduction or not. Unlike machines, whose governing functions are embedded by human designers, organisms are self-governing. The autopoietic definition of life resembles the physiological definition but emphasizes life’s maintenance of its own identity, its informational closure, its cybernetic self-relatedness, and its ability to make more of itself. Autopoiesis refers to self-producing, self-maintaining, self-repairing, and self-relational aspects of living systems. Living beings maintain their form by the continuous interchange and flow of chemical components. Cellular autopoietic systems are bounded by a dynamic material made by the system itself. In life on Earth the limiting material is lipidmembrane studded with transport proteins fabricated by the incessantly active cell. A source of usable energy flows to all living or autopoietic systems—either light in the visible or near-visible spectrum or specific organic carbon or other chemicals such as hydrogen,hydrogen sulfide, or ammonia. Energy sources that have never been adequate to maintain autopoiesis on Earth include heat, sound waves, and electromagnetic radiation outside the visible or near-visible spectrum.
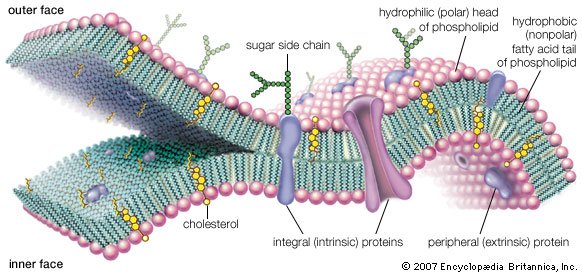
Molecular view of the cell membrane
Intrinsic proteins penetrate and bind tightly to the lipid bilayer, which is made up largely of phospholipids and cholesterol and which typically is between 4 and 10 nanometers (nm; 1 nm = 10−9 metre) in thickness. Extrinsic proteins are loosely bound to the hydrophilic (polar) surfaces, which face the watery medium both inside and outside the cell. Some intrinsic proteins present sugar side chains on the cell's outer surface.
One of the difficulties in defining life is that the only example is life found on the third planet from the Sun. On Earth all life’s autopoietic systems require a supply of water in its liquid state for self-maintenance of their parts. Taken together, all transformations that underlie autopoiesis require six elements: carbon, nitrogen, hydrogen, oxygen, phosphorus, and sulfur. The chemical components of all living entities are fashioned primarily from these elements.
The smallest autopoietic system on Earth is a living bacterial cell. (Viruses, plasmids, and other replicating molecules cannot, even in principle, behave as an autopoietic system; no matter how much food, liquid water, and serviceable energy they are provided, they still require cells for their continuity and duplication.) Some cells, such as Carsonella ruddii, have fewer than 200 genes and proteins, but they, like organelles and viruses, are not autopoietic, since they must be inside an autopoietic system (living cell) to metabolize and survive. No self-bounded autopoietic system smaller than a cell with at least 450 proteins and the genes that code for these proteins has ever been described. Larger than bacteria are other autopoietic systems of intermediate size such as protists, fungal spores, mules and other individual mammals, and plants such as oak trees or poppies. Autopoietic entities at even larger levels include ecosystems such as coral reefs,prairies, or ponds. The maximal or largest single autopoietic system known is referred to as “Gaia,” named by English atmospheric scientist James E. Lovelock for Gaea, the ancient Greek personification of Earth. Gaia is basically a closed thermodynamic system because there is little interchange of matter with the extraterrestrial environment. There is evidence that the global, Gaian system of life shows organism-like properties, such as regulation of atmospheric chemistry, global mean temperature, and oceanic salinity over multimillion-year time spans. Such regulation may be understood as part of life’s organization as a complex and cyclical open thermodynamic system.
Life On Earth
Life On Earth
Life On Earth (B)
The existence of diverse definitions of life, as detailed in the previous section, surely means that life is complex and difficult to briefly define. A scientific understanding of living systems has existed since the second half of the 19th century. But the diversity of definitions and lack of consensus among professionals suggest something else as well. As detailed in this section, all organisms on Earth are extremely closely related, despite superficial differences. The fundamental pattern, both in form and in matter, of all life on Earth is essentially identical. Also, as noted in this section, this identity implies that all organisms on Earth are evolved from a single instance of the origin of life. To generalize from a single example is difficult, especially when the example itself is changing, growing, and evolving. In this respect the biologist is fundamentally handicapped, as compared with, say, the chemist, physicist, geologist, or meteorologist, each of whom can now study aspects of his discipline beyond Earth. If truly only one sort of life on Earth exists, then perspective is lacking in a most fundamental way. On the other hand, the historical continuity of all life-forms means that ancient life, perhaps even the origins of life, may be glimpsed by studying modern cells.
Over hundreds of millions of years, life spread through the seas and over Earth's surface. The first life-forms were small and simple. Later forms were more complicated and diverse.
The biosphere
The biosphere (B)
Reconnaissance missions to the planets of the inner solar system have revealed stark and barren landscapes. From the heavily cratered and atmosphereless surfaces of both Mercury and the Moon to the hot sulfurous fogs of Venus and the dusty, windswept surface of Mars, no sign of life is apparent anywhere. The biosphere, by definition the place where all Earth’s life dwells, is a delight with its green, wet contrast. Austrian geologist Eduard Suess invented the term biosphere to match the other envelopes of the planet: the atmosphere of gas; the hydrosphere of oceans,lakes,rivers,springs and other waters; and the lithosphere, or the solid rock surface of the outer portion of Earth. Yet it was the great Russian crystallographer and mineralogist Vladimir I. Vernadsky who brought the term into common parlance with his book of the same name. In The Biosphere (1926) Vernadsky outlines his view of life as a major geological force. Living matter, Vernadsky contends, erodes, levels, transports, and chemically transforms surface rocks, minerals, and other features of Earth. If the biosphere is the place where life is found, the biota (or the biomass as a whole) is the sum of all living forms: flora,fauna, and microbiota.
The planet Earth.
During the second half of the 20th century, study of the deep sea, the upper atmosphere, the Antarctic dry deserts, newly opened caves, sulfurous tunnels, and granitic rocks showed that Earth’s surface is vigorously inhabited in places that were unknown to Vernadsky and his colleagues. Vernadsky’s international school of thought ushered in the field of “biogeochemistry,” and chemists and geologists were recruited to consider life as a planetary phenomenon. But not until giant, mouthless, red-gilled tube worms were videographed in the late 1970s and ’80s did the extent and the weirdness of Earth’s biota begin to be fathomed. Entire large ecosystems were recognized on the ocean’s bottom that live not by the usual plantphotosynthesis but rather by chemolithoautotrophy, a kind of metabolism in which organisms make food from carbon dioxide using energy from the oxidation of sulfide, methane, or other inorganic compounds. These discoveries have led to a deeper understanding of life’s varied modes of nutrition and sources of energy. Bacterial symbionts living in the tissues of some polychaeteworms (alvinellids) or pogonophora (such as Riftia pacytila) provide the animals with their total nutritional needs. The submarine ecosystems supported by bacteria thrive along the worldwide rift zones that extend along the borders of huge continental plates at the Mid-Atlantic Ridge, on the East Pacific Rise, at 21° north of the Equator off the coast of Baja California, Mex., and at a dozen other newly studied sites. By the beginning of the 21st century it had become abundantly clear that many life-forms and ecosystems remained unknown or under-studied. Those in the Siberian tundra, in the thickly forested portions of the Amazon River valley and its tributaries, at the tops of remote mountains and inside granitic rocks in temperate zones, and in the centre of Africa remain as inaccessible to most naturalists as they have been throughout history. The easily accessed woodlands and fields of well-lit land surfaces are another story.
On land, 24 percent of the productivity of organic carbonbiomass generated by plants is directly controlled by burgeoning populations of one species, humans. As Vernadsky noted, life in general and human life in particular tend to accelerate the number of materials and the rate of flow of these materials through the biosphere, the place where all life exists—so far.
Vernadsky anticipated new discoveries of life inside hot springs and granitic rock. Although he qualified this statement by asserting that it would not hold for temporary, abnormal circumstances, such as during a lava flow or a volcanic eruption, he wrote,
“Thus far, we have seen that the biosphere, by structure, composition, and physical makeup, is completely enclosed by the domain of life, which has so adapted itself to biospheric conditions that there is no place [on Earth] in which it is unable to manifest itself in one way or another.”
Although much is not known about life in the depths of the rocks and the sea, determination of the total range and mass (biomass) of the biota, the sum of all life in the biosphere, is a reasonable scientific goal.
Chemistry of life
Chemistry of life (B)
Human beings, like mammals in general, are ambulatory collections of some 1014cells. Human cells are in all fundamental respects the same as those that make up the other animals. Each cell typically consists of one central, spherical nucleus and another heterogeneous region, the cytoplasm. (Only bacterial cells lack nuclei; those of plants, fungi, and all other organisms contain one or more nuclei.) A living nucleated cell, a marvel of detailed and complex architecture, appears frenetic with activity when seen through a microscope. On a deeper chemical level, it is known that life’s large molecules, the proteins and nucleic acids, are synthesized at a very fast rate. Enzymes, which speed up chemical reactions, are all proteins, but by no means are all proteins enzymes. An enzyme catalyzes the synthesis of more than 1,000 other molecules per second. The total mass of a metabolizing bacterial cell can be synthesized in 20 minutes. The information content of a small cell has been estimated as about 1010bits, comparable to about 106 (or one million) pages of the print version of the Encyclopædia Britannica. Although some feel debased by the implication that people are “nothing more” than a frenetic collection of interacting molecules, others are thrilled with the power of science to reveal the inner workings of the chemistry of life. The spectacular success of biochemistry and molecular biology in the 20th century suggests that laws of biology are derived from the interaction of atoms, thermodynamic principles, and life’s chemistry, which has persisted with faithful continuity since its origin some 3.7 billion to 3.5 billion years ago.
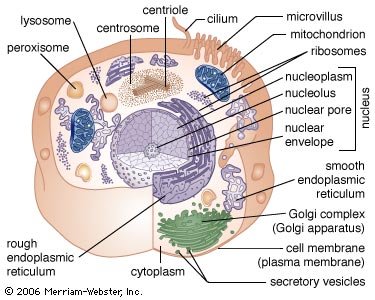
Animal cell
Principal structures of an animal cell
Cytoplasm surrounds the cell's specialized structures, or organelles. Ribosomes, the sites of protein synthesis, are found free in the cytoplasm or attached to the endoplasmic reticulum, through which materials are transported throughout the cell. Energy needed by the cell is released by the mitochondria. The Golgi complex, stacks of flattened sacs, processes and packages materials to be released from the cell in secretory vesicles. Digestive enzymes are contained in lysosomes. Peroxisomes contain enzymes that detoxify dangerous substances. The centrosome contains the centrioles, which play a role in cell division. The microvilli are fingerlike extensions found on certain cells. Cilia, hairlike structures that extend from the surface of many cells, can create movement of surrounding fluid. The nuclear envelope, a double membrane surrounding the nucleus, contains pores that control the movement of substances into and out of the nucleoplasm. Chromatin, a combination of DNA and proteins that coil into chromosomes, makes up much of the nucleoplasm. The dense nucleolus is the site of ribosome production.
DNA, RNA, and protein
DNA, RNA, and protein (B)
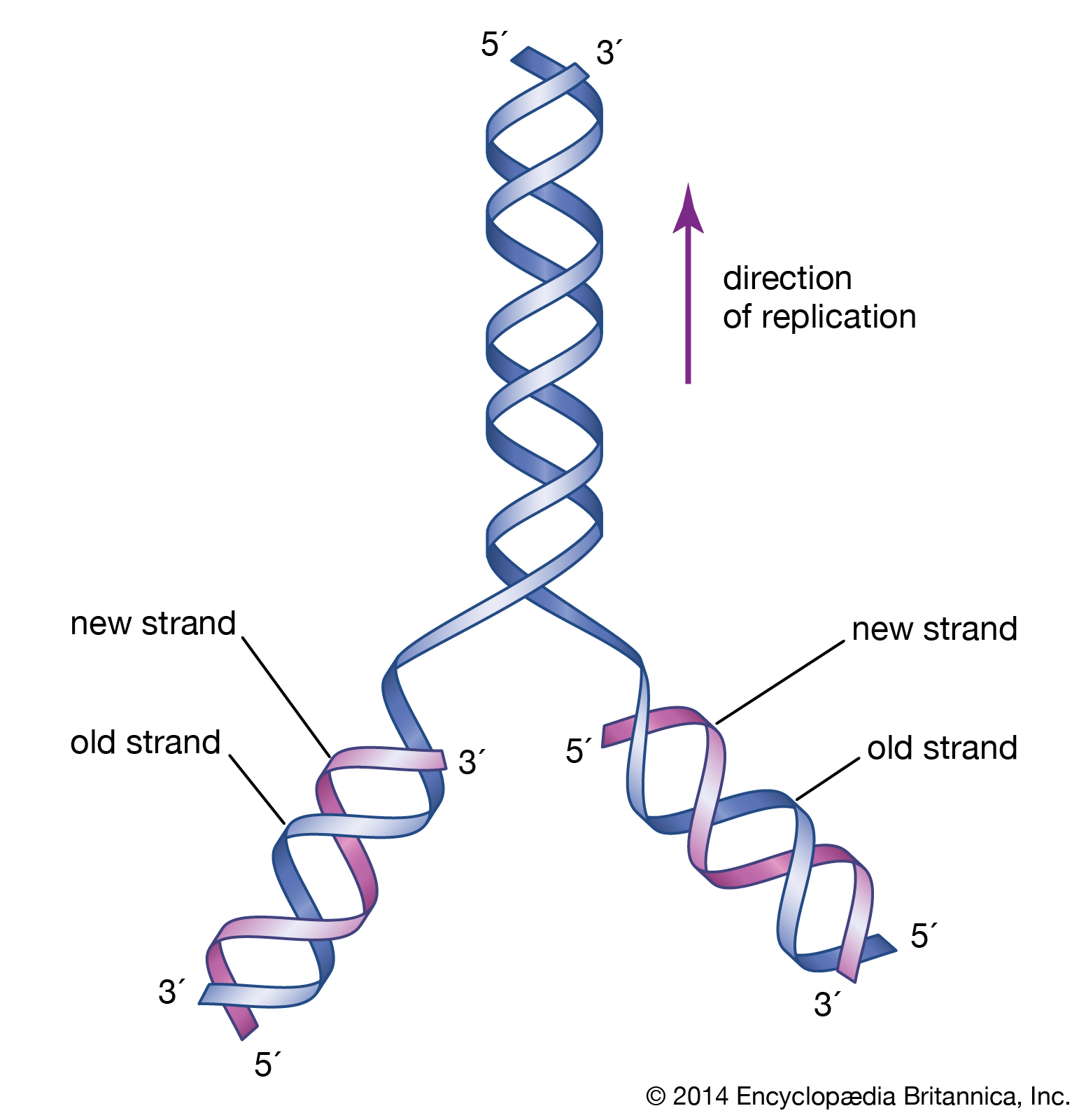
The specific carrier of the genetic information in all organisms is the nucleic acid known as DNA, short for deoxyribonucleic acid. DNA is a double helix, two molecular coils wrapped around each other and chemically bound one to another by bonds connecting adjacent bases. Each long ladderlike DNA helix has a backbone that consists of a sequence of alternating sugars and phosphates. Attached to each sugar is a “base” consisting of the nitrogen-containing compound adenine, guanine, ctyosine, or thymine. Each sugar-phosphate-base “rung” is called a nucleotide. A very significant one-to-one pairing between bases occurs that ensures the connection of adjacent helices. Once the sequence of bases along one helix (half the ladder) has been specified, the sequence along the other half is also specified. The specificity of base pairing plays a key role in the replication of the DNA molecule. Each helix makes an identical copy of the other from molecular building blocks in the cell. These nucleic acid replication events are mediated by enzymes called DNA polymerases. With the aid of enzymes, DNA can be produced in the laboratory.
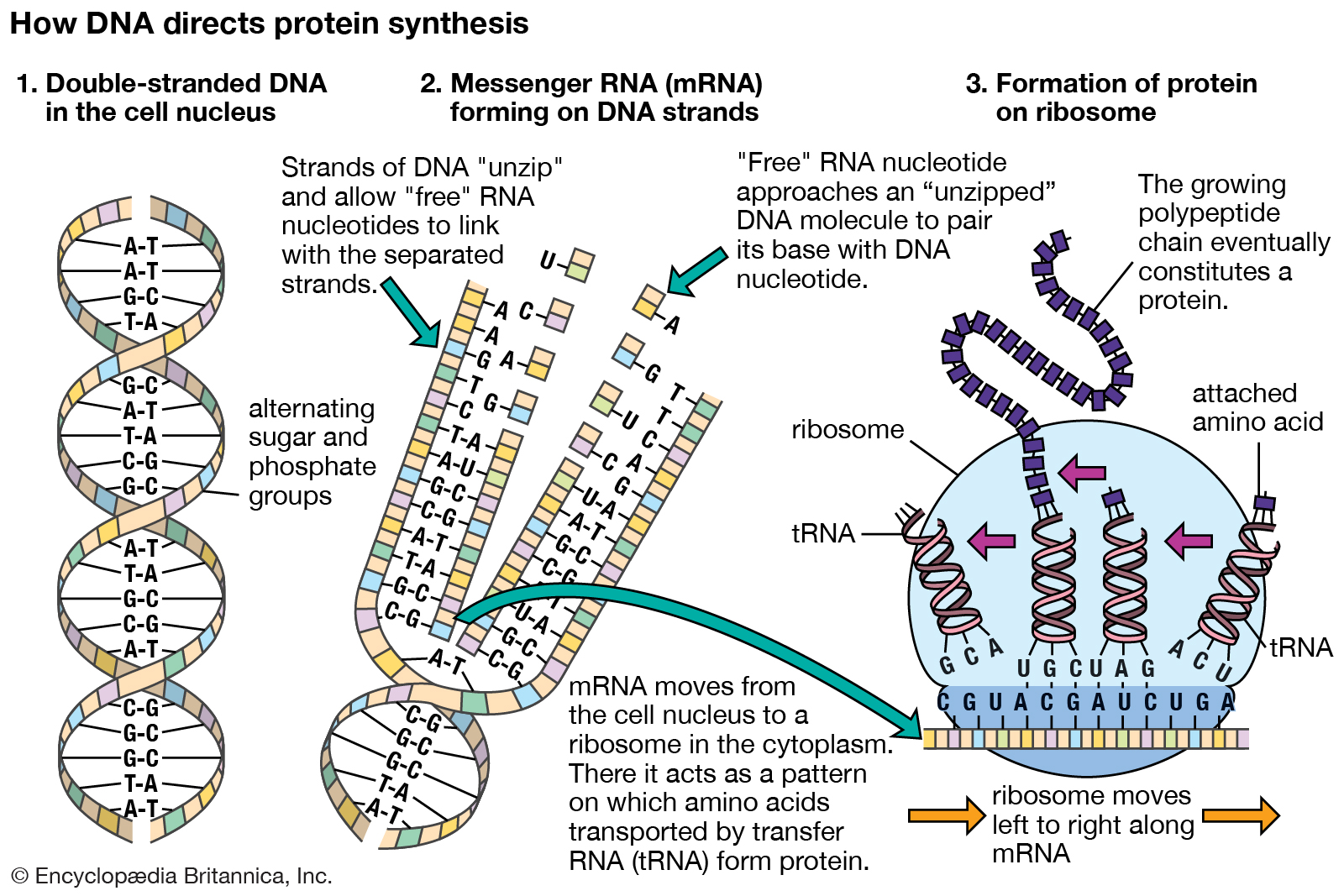
DNA and protein synthesis
DNA in the cell nucleus carries a genetic code, which consists of sequences of adenine (A), thymine (T), guanine (G), and cytosine (C) (Figure 1). RNA, which contains uracil (U) instead of thymine, carries the code to protein-making sites in the cell. To make RNA, DNA pairs its bases with those of the “free” nucleotides (Figure 2). Messenger RNA (mRNA) then travels to the ribosomes in the cell cytoplasm, where protein synthesis occurs (Figure 3). The base triplets of transfer RNA (tRNA) pair with those of mRNA and at the same time deposit their amino acids on the growing protein chain. Finally, the synthesized protein is released to perform its task in the cell or elsewhere in the body.
The cell, whether bacterial or nucleated, is the minimal unit of life. Many of the fundamental properties of cells are a function of their nucleic acids, their proteins, and the interactions among these molecules bounded by active membranes. Within the nuclear regions of cells is a mélange of twisted and interwoven fine threads, the chromosomes. Chromosomes by weight are composed of 50–60 percent protein and 40–50 percent DNA. During cell division, in all cells but those of bacteria (and some ancestral protists), the chromosomes display an elegantly choreographed movement, separating so that each offspring of the original cell receives an equal complement of chromosomal material. This pattern of segregation corresponds in all details to the theoretically predicted pattern of segregation of the genetic material implied by the fundamental genetic laws (seeheredity). The chromosome combination of the DNA and the proteins (histone or protamine) is called nucleoprotein. The DNA stripped of its protein is known to carry genetic information and to determine details of proteins produced in the cytoplasm of cells; the proteins in nucleoprotein regulate the shape, behaviour, and activities of the chromosomes themselves.
The other major nucleic acid is ribonucleic acid (RNA). Its five-carbon sugar is slightly different from that of DNA. Thymine, one of the four bases that make up DNA, is replaced in RNA by the base uracil. RNA appears in a single-stranded form rather than a double. Proteins (including all enzymes), DNA, and RNA have a curiously interconnected relation that appears ubiquitous in all organisms on Earth today. RNA, which can replicate itself as well as code for protein, may be older than DNA in the history of life.
Chemistry in common
Chemistry in common (B)
The genetic code was first broken in the 1960s. Three consecutive nucleotides (base-sugar-phosphate rungs) are the code for one amino acid of a protein molecule. By controlling the synthesis of enzymes, DNA controls the functioning of the cell. Of the four different bases taken three at a time, there are 43, or 64, possible combinations. The meaning of each of these combinations, or codons, is known. Most of them represent one of the 20 particular amino acids found in protein. A few of them represent punctuation marks—for example, instructions to start or stop protein synthesis. Some of the code is called degenerate. This term refers to the fact that more than one nucleotide triplet may specify a given amino acid. This nucleic acid–protein interaction underlies living processes in all organisms on Earth today. Not only are these processes the same in all cells of all organisms, but even the particular “dictionary” that is used for the transcription of DNA information into protein information is essentially the same. Moreover, this code has various chemical advantages over other conceivable codes. The complexity, ubiquity, and advantages argue that the present interactions among proteins and nucleic acids are themselves the product of a long evolutionary history. They must interact as a single reproductive, autopoietic system that has not failed since its origin. The complexity reflects time during which natural selection could accrue variations; the ubiquity reflects a reproductive diaspora from a common genetic source; and the advantages, such as the limited number of codons, may reflect an elegance born of use. DNA’s “staircase” structure allows for easy increases in length. At the time of the origin of life, this complex replication and transcription apparatus could not have been in operation. A fundamental problem in the origin of life is the question of the origin and early evolution of the genetic code.
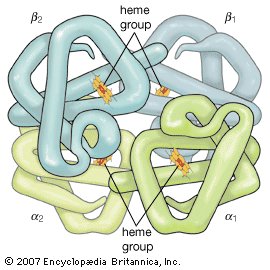
The hemoglobin tetramer
Two αβ dimers combine to form the complete hemoglobin molecule. Each heme group contains a central iron atom, which is available to bind a molecule of oxygen. The α1β2 region is the area where the α1 subunit interacts with the β2 subunit.
Many other commonalities exist among organisms on Earth. Only one class of molecules stores energy for biological processes until the cell has use for it; these molecules are all nucleotide phosphates. The most common example is adenosine triphosphate (ATP). For the very different function of energy storage, a molecule identical to one of the building blocks of the nucleic acids (both DNA and RNA) is employed. Metabolically ubiquitous molecules—flavin adenine dinucleotide (FAD) and coenzyme A—include subunits similar to the nucleotide phosphates. Nitrogen-rich ring compounds, called porphyrins, represent another category of molecules; they are smaller than proteins and nucleic acids and common in cells. Porphyrins are the chemical bases of the heme in hemoglobin, which carries oxygen molecules through the bloodstream of animals and the nodules of leguminous plants. Chlorophyll, the fundamental molecule mediating light absorption during photosynthesis in plants and bacteria, is also a porphyrin. In all organisms on Earth, many biological molecules have the same “handedness” (these molecules can have both “left-” and “right-handed” forms that are mirror images of each other; see belowThe earliest living systems). Of the billions of possible organic compounds, fewer than 1,500 are employed by contemporary life on Earth, and these are constructed from fewer than 50 simple molecular building blocks.
Paramecium caudatum (highly magnified).
Besides chemistry, cellular life has certain supramolecular structures in common. Organisms as diverse as single-celled paramecia and multicellular pandas (in their sperm tails), for example, possess little whiplike appendages called cilia (or flagella, a term that is also used for completely unrelated bacterial structures; the correct generic term is undulipodia). These “moving cell hairs” are used to propel the cells through liquid. The cross-sectional structure of undulipodia shows nine pairs of peripheral tubes and one pair of internal tubes made of proteins called microtubules. These tubules are made of the same protein as that in the mitotic spindle, the structure to which chromosomes are attached in cell division. There is no immediately obvious selective advantage of the 9:1 ratio. Rather, these commonalities indicate that a few functional patterns based on common chemistry are used over and over again by the living cell. The underlying relations, particularly where no obvious selective advantage exists, show all organisms on Earth are related and descended from a very few common cellular ancestors—or perhaps one.
Modes of nutrition and energy generation
Modes of nutrition and energy generation (B) Chemical bonds that make up the compounds of living organisms have a certain probability of spontaneous breakage. Accordingly, mechanisms exist that repair this damage or replace the broken molecules. Furthermore, the meticulous control that cells exercise over their internal activities requires the continued synthesis of new molecules. Processes of synthesis and breakdown of the molecular components of cells are collectively termed metabolism. For synthesis to keep ahead of the thermodynamic tendencies toward breakdown, energy must be continuously supplied to the living system.
Energy, carbon, and electrons
Energy, carbon, and electrons (B)
Organisms acquire energy by two general methods: by light or by chemical oxidation. Productive organisms, called autotrophs, convert light or chemicals into energy-rich organic compounds beginning with energy-poor carbon dioxide (CO2). These autotrophs provide energy for the other organisms, the heterotrophs. Heterotrophs are organisms that acquire their energy by the controlled breakdown of preexisting organic molecules, or food. Human beings, like most other animals, fungi, protists, and bacteria, are heterotrophs.
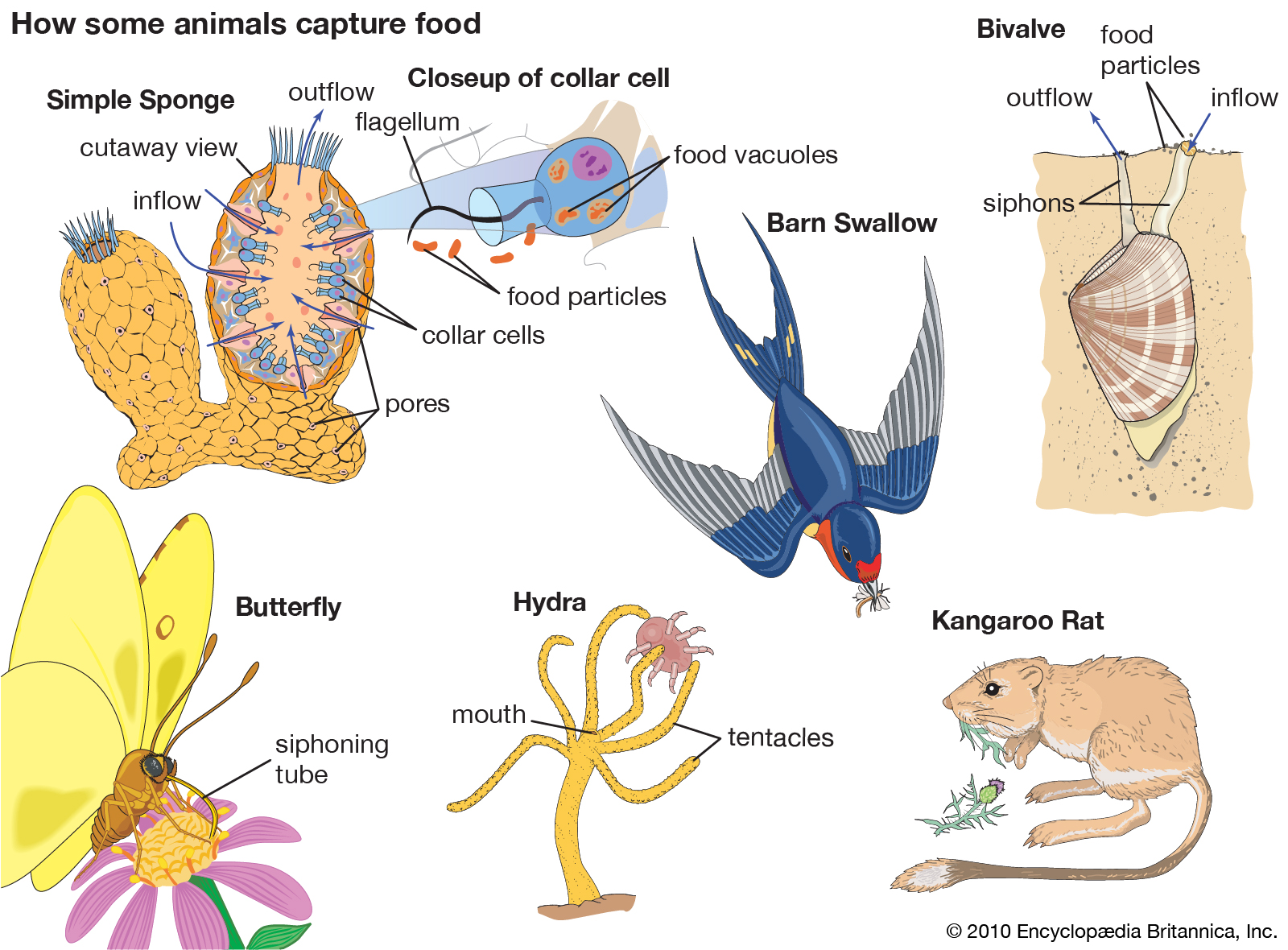
Food capture: animals
The process of food capture in different animal species.
Autotrophic organisms are often primary producers in their ecosystems. They acquire their useful free energy from sources other than food: either from the energy of sunlight (photoautotrophs) or from oxidative chemical reactions (chemoautotrophs). The latter mode of metabolism refers to life-forms that use inorganic materials (ammonia [NH3], methane [CH4], or hydrogen sulfide [H2S]) combined with oxygen to generate their energy. Only some bacteria are capable of obtaining energy by “burning” inorganic chemicals.
Green plants are typical photoautotrophs. Plants absorb sunlight to generate ATP and to disassociate water into oxygen and hydrogen. To break down the water molecule, H2O, into hydrogen and oxygen requires much energy. The hydrogen from water is then combined in the “dark reactions” with carbon dioxide, CO2. The result is the production of such energy-rich organic molecules as sugars, amino acids, and nucleotides. The oxygen becomes the gas O2, which is released as waste back into the atmosphere. Animals, which are strictly heterotrophs, cannot live on carbon dioxide, sunlight, and water with a few salts like plants do. They must breathe in the atmospheric oxygen. Animals combine oxygen chemically with hydrogen atoms that they remove from their food—that is, from organic materials such as sugar, protein, and amino acids. Animals release water as a waste product from the oxygen respiration. Animals, like all heterotrophs, use organic materials as their sole source of carbon. This conversion of carbon provides an example of an aspect of an ecological cycle in which a required element flows through different types of organisms as it changes its oxidation state from CO2 to (CH2O)n and back to CO2.
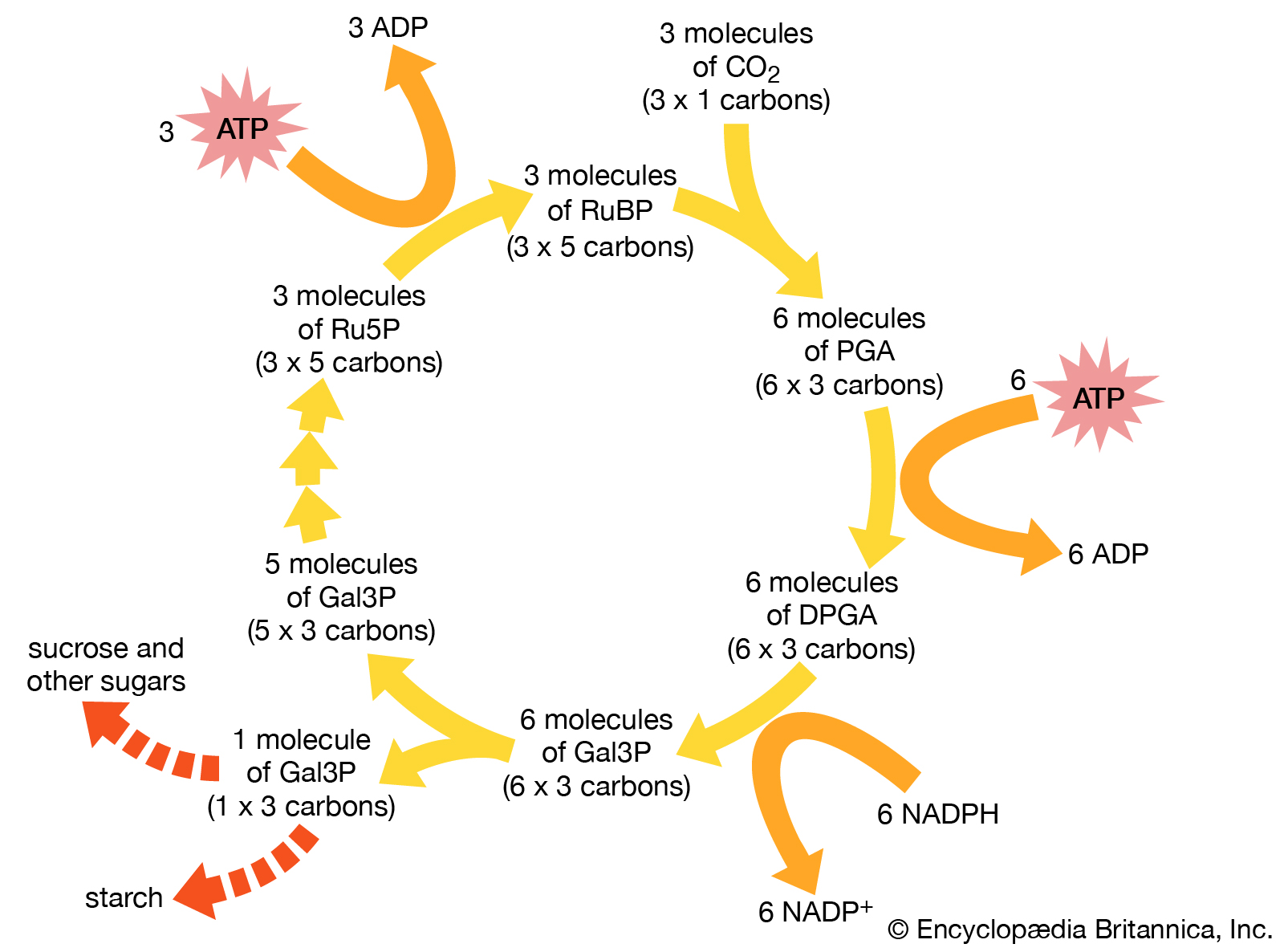
Calvin cycle
Pathway of carbon dioxide fixation and reduction in photosynthesis, the Calvin cycle. The diagram represents one complete turn of the cycle, with the net production of one molecule of glyceraldehyde-3-phosphate (Gal3P). This three-carbon sugar phosphate usually is converted to either sucrose or starch.
Metabolic cycles in general—the extraction by organisms of useful energy and food molecules from environmental source material—can be described in terms of oxidation-reduction reactions. In the case of oxygen respiration, oxygen molecules from the air accept electrons ultimately from glucose or amino acids. The oxygen, which has a great affinity for electrons, is called an electron acceptor, whereas the glucose, or other sugar or organic molecules, is an electron donor. Animal respiration is the prototype of oxidation-reduction reactions, but certainly not all oxidation-reduction reactions (or “redox reactions,” as they are often called) involve oxygen. Many other inorganic compounds are respired, or “breathed,” at the cell level. Biological electron acceptors besides oxygen include nitrate,nitrite,sulfate,carbonate, elemental sulfur, and methanol. Biological electron donors (other than sugar and amino acids) include hydrogen, nitrogen compounds (as ammonia, nitrite), sulfide, and methane. For acceptor-donor transformations to be available to chemoautotrophs and heterotrophs over sustained periods of time, ecological cycles are required. For geologically short periods of time, organisms may live off a finite supply of material; however, for any long-term continuance of life, a dynamic cycling of matter involving complementary types of organisms must prevail. If life exists on other planets, the requisite elements and liquid water must cycle. A search for such transformations provides one method of detecting extraterrestrial life.
In addition to energy, all forms of life require carbon sources. Autotrophic organisms (chemosynthetic and photosynthetic bacteria, algae, and plants) derive this essential element from carbon dioxide. Heterotrophs use preformed organic compounds as their source of carbon. Among autotrophs many types of cells do not depend on light to generate ATP; those that do without light are the chemoautotrophic bacteria, including the methanogens, ammonia oxidizers, sulfide oxidizers, hydrogen oxidizers, and a few obscure others. Indeed, at least five metabolic pathways entirely different from each other have evolved to use carbon dioxide gas. One is the oxygenic pathway described above, which is used by plants, algae, and cyanobacteria: the Calvin-Benson dark reactions. Other, more obscure pathways include phosphoenolpyruvate (PEP), succinate, and methanogen pathways. They all need to bring energy-poor carbon dioxide into the energy-rich carbon-hydrogen compound metabolism of organisms. All life on Earth depends on these autotrophic reactions that begin with carbon dioxide or its equivalent. Equivalents as carbon sources in autotrophic metabolism include the carbonate ion, bicarbonate ion, and carbon monoxide. As usual, with respect to metabolic variation and virtuosity, the bacterial repertoire is vastly more diverse than that of eukaryotes—that is, plants, animals, and other organisms composed of cells with nuclei. In general, nucleated organisms, eukaryotes, are either photolithoautotrophs (i.e., algae and plants) that derive energy from light or minerals or chemo-organoheterotrophs (animals, fungi, and most protists) that derive energy and carbon from preformed organic compounds (food).
All ATP biological electron-transfer reactions lead to the net production of ATP molecules. Two of the three phosphates (PO4) of this molecule are held by energy-rich bonds sufficiently stable to survive for long periods of time in the cell but not so strong that the cell cannot tap these bonds for energy when needed. ATP and similar molecules (such as guanosine triphosphate [GTP]) have a five-carbon sugar and three phosphates. As far as is known, such molecules are the general and unique energy currency of living systems on Earth.
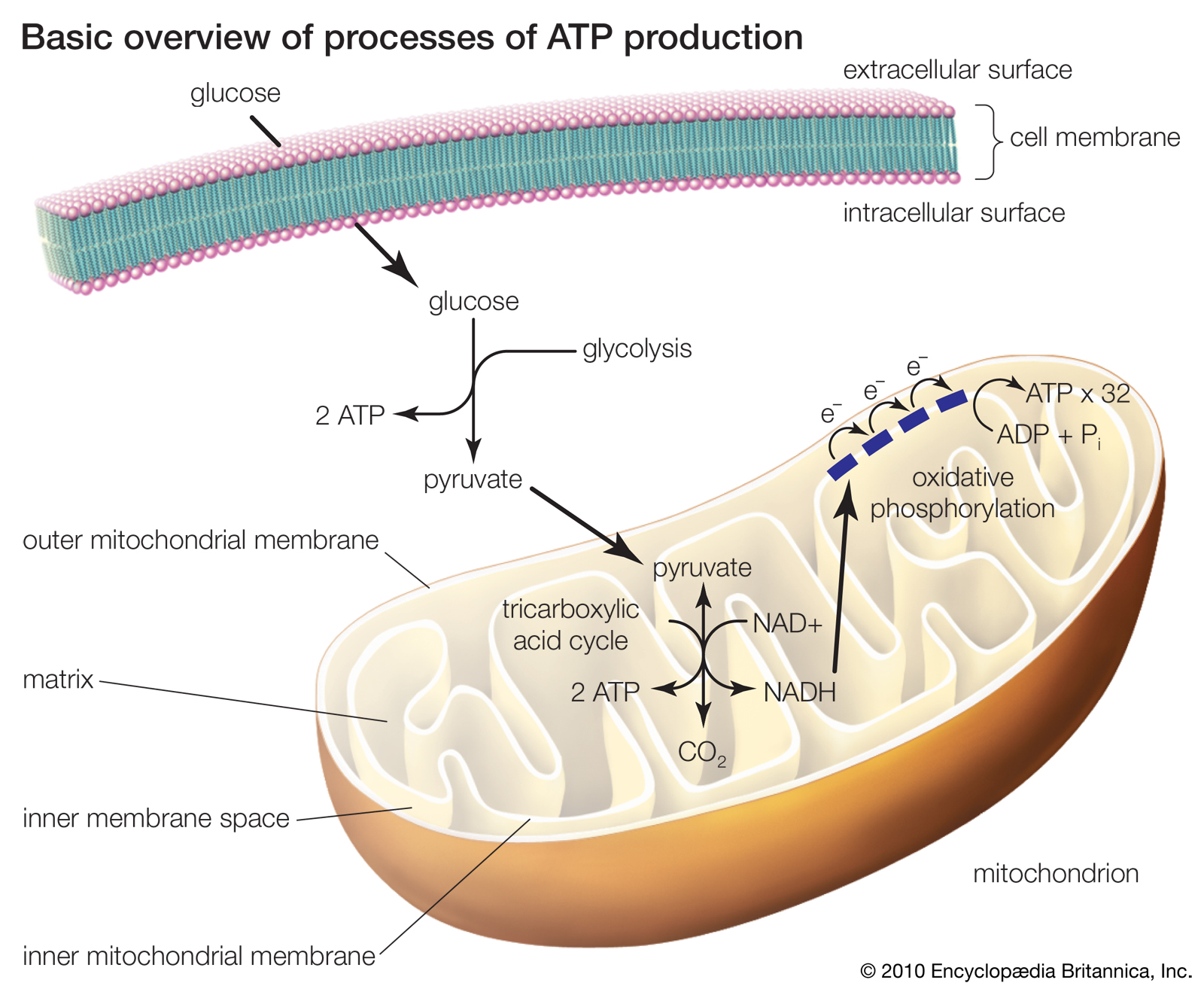
Basic overview of processes of ATP production
The three processes of ATP production include glycolysis, the tricarboxylic acid cycle, and oxidative phosphorylation. In eukaryotic cells the latter two processes occur within mitochondria. Electrons that are passed through the electron transport chain ultimately generate free energy capable of driving the phosphorylation of ADP.
No metabolic process occurs in a single step. The ordinary six-carbon sugar, glucose, does not oxidize to carbon dioxide and water in living cells in the same way that glucose in air burns. Any release of energy by burning would be too sudden and too concentrated in a small volume to happen safely inside the tiny cell. Instead, glucose is broken down at ambient (i.e., relatively cool) temperatures by a series of successive and coordinated steps. Each step is mediated by a particular and specific enzyme. In most cells that metabolize glucose, the sugar first breaks down in a set of steps that occur in the absence of oxygen. The total number of such steps in plants, animals, fungi, and protists (see belowHypotheses of origins) is about 11. Other organisms, primarily bacteria and obscure protists and fungi, are anaerobes: they do not utilize molecular oxygen in their metabolism. In anaerobes, glucose metabolism stops at compounds such as ethanol or lactic acid.Aerobic organisms, including all animals, carry the oxidation of glucose farther. They rapidly use anaerobic glucose breakdown products such as lactic acid, ethanol, or acetate with Krebs-cycle intermediates in the mitochondria.Aerobic oxidation of glucose requires an additional 60 enzyme-catalyzed steps. The anaerobic breakdown of glucose uses enzymes suspended freely in solution in the cells. The aerobic steps occur on enzymes localized in mitochondria, the “power packs” of cells where oxygen gas is used to make the energy compound ATP. The complete aerobic breakdown of sugar to carbon dioxide and water is about 10 times more efficient than the anaerobic in that 10 times as many ATP molecules are produced. (Seemetabolism.)
Energy made available to cells in the form of ATP is used in a variety of ways—for example, for motility. When an amoeba extends pseudopods or when a person walks, ATP molecules are tapped for their energy-rich phosphate bonds. ATP molecules are used for the synthesis of proteins that all cells require in their growth and division, amino acids, and five-carbon sugars of nucleic acids. Each synthetic process is controlled and enzymatically mediated. Each starts from an organic building-block compound available to the cell as food. The amino acid L-leucine, for example, is produced from pyruvic acid, which is itself the product of the anaerobic breakdown of glucose. Synthesis of L-leucine from pyruvic acid involves eight enzyme-mediated steps with an addition of acetic acid and water.
These exquisitely interlocked and controlled metabolic steps are not performed in a diffuse manner helter-skelter in the cell. Rather, a marvelously architectured cellular interior displays specialized regions visible at the electron-microscopic level. Particular chemical reactions are performed in association with specific structures. In aerobic eukaryotes the mitochondrion with its intricate cristate membrane (the folds in the membrane are called cristae) provides the site of pyruvate, acetate, and lactate metabolism. These molecules are transformed and passed on from one enzyme to another as through a conveyor belt in a factory. Similarly, in those eukaryotes capable of oxygenic photosynthesis (algae, plants), photosynthesis occurs only in an organelle (a cell part) called a chloroplast. Chlorophyll, carotenoids, and other pigments that absorb visible light, as well as the detailed enzymatic apparatus for the photosynthetic process, reside there.
Chloroplasts and mitochondria contain DNA. Moreover, this DNA has a sequence distribution that differs entirely from that of the nucleus and greatly resembles that of free-living photosynthetic and oxygen-respiring bacteria. The best explanation for these facts is that the ancestors to the oxygen-releasing chloroplasts and oxygen-respiring mitochondria were once free-living bacteria.
All life is composed of cells of one of two types: prokaryotes (those that lack a nucleus) or eukaryotes (those with a nucleus). Even in one-celled organisms this distinction is very clear.
All bacteria are prokaryotic, even though many, probably most, are multicelled in nature. The only other single-celled organisms that exist are fungi (one-celled fungi are called yeasts). All nucleated organisms (cells with nuclei and chromosomes in their cells) that are not animals, fungi, or plants are Protista. This huge group includes the unicellular or few-celled protists and their multicellular descendants. The large kingdom of Protista has 250,000 estimated species alive today. Some are very large, such as red algae and the kelp Macrocystis. One-celled protists include the familiar amoebas, paramecia, and euglenas as well as 50,000 less-familiar types. Scientifically speaking, no such thing as a one-celled animal exists. All animals and plants are by definition multicellular, since they all develop from multicellular embryos. Accordingly, all “protozoans” are now classified as single-celled Protista, not animals. Nor are there any one-celled plants. Organisms formerly called one-celled plants are algae and, as such, are now classified with Protista as well. If a mature organism is determined to be one-celled, then it must be either a bacterium (prokaryotic) or a fungus or protist (eukaryotic). All animals and plants develop from embryos that by definition combine two complementary sets of chromosomes (i.e., they are diploids at some stage in their development). They are all multicellular eukaryotes. But though there are no one-celled plants or animals, there are indeed myriad many-celled protists. Multicellularity evolved not only in the ancestors to the plants and the animals but also in the bacteria, the protists, and the fungi.
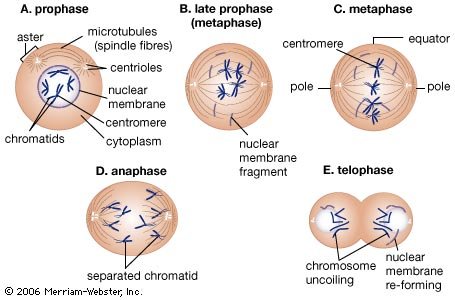
All eukaryotic cells undergo some form of mitosis, a sequence of cell division events that occurs after chromosomal DNA protein replication. Mitosis ensures that chromosomal DNA and protein are equally distributed to the offspring cells. Mitosis is the most distinctive activity of eukaryotic cells, which have nucleoprotein chromosomes in their nuclei and a membrane that separates the nucleus from the cytoplasm. In mitosis, mitochondria, which are usually present in the cytoplasm as well as in the chloroplasts of algae and plants, are smoothly distributed with the chromosomes to offspring cells. The Golgi apparatus and endoplasmic reticulum (ER), an intricately convoluted structure, serve to anchor many cytoplasmic enzymes excluded from mitochondria or chloroplasts. They also divide and are distributed in mitosis.
Stages of mitosis. A. Prophase. Replicated chromosomes, consisting of two daughter strands (chromatids) attached by a centromere, coil and contract. Two pairs of specialized organelles (centrioles) begin to move apart, forming a bridge of hollow protein cylinders known as microtubules (spindle fibres) between them. Microtubules also extend in a radial array (aster) from the centrioles to the poles of the cell. B. Late prophase. As the centrioles move apart, the nuclear membrane breaks down and microtubules extend from each centromere to opposite sides or poles of the cell. C. Metaphase. The centromeres align in a plane midway between the poles known as the equator, or metaphase plate. During late metaphase, each centromere divides into two, freeing sister chromatids from each other. D. Anaphase. Sister chromatids are drawn to opposite ends as centromeric microtubules shorten and polar microtubules lengthen, causing the poles to move farther apart. E. Telophase. Chromosomes uncoil, microtubules disappear, and the nuclear membrane re-forms around each set of daughter chromosomes. The cytoplasm begins to pinch in to create two daughter cells.
Nuclei, chromosomes, mitochondria, chloroplasts, ER, and nuclear membranes are all absent in prokaryotes. Prokaryotic cells, which include all the cyanobacteria (formerly called blue-green algae), are bacteria in every way. Division is nonmitotic in all prokaryotes. Bacteria lack nucleoprotein and a nuclear membrane, and, when chromosome stain is applied, only fuzz or nothing is seen. Whereas all eukaryotic cells have more than one chromosome and sometimes over a thousand, the genes of prokaryotic cells are organized into a single “chromoneme” or “genophore.” (The term bacterial chromosome, while still in use, is, technically speaking, inaccurate.) The genes may or may not be concentrated enough to be seen, but in any case bacterial DNA floats freely in the cytoplasm. Prokaryote cell organization is less complex than that of eukaryotes. The basic question of the evolution of prokaryotes into eukaryotes—often rated the second major evolutionary mystery, after the origin of life—is thought to involve a complex series of partnerships in which distinct strains of bacteria entered each others’ bodies, merged symbiotically, and traded genes.
Since multicellularity evolved independently in every major group of microorganisms, the blurred distinction between single-celled and many-celled organisms has become obsolete. The protists are divisible into about 35 unambiguous groups called phyla. They provide many examples of biological principles—including the prevalence of independent trends toward multicellularity. One illustration involves cellular slime molds. These heterotrophs undergo an extraordinary sequence of events during their life history. The story begins with single cells, indistinguishable from common amoebas. When starved, they begin to swarm. Soon they combine into a slimy mass of many nucleated amoeba cells called a pseudoplasmodium. The pseudoplasmodium in turn forms a sluglike multicellular creature resembling a mollusk that has escaped from its shell. This slug, which is entirely multicellular, migrates and then stops and develops into a stalk structure called a sorocarp that bears amoeba cysts on top. The cysts were called “spores.” Some have cellulose cell walls similar to those of plants. The cysts, which are encased amoebas (just like other amoeba cysts), germinate in turn—when water and food again become abundant—into new amoebas. The released amoebas extend their pseudopods, and, as individuals again, they migrate to feed. The life history repeats with swarms of migrating amoebas, slugs, stalks, and finally clusters of amoeba cysts on top as wet, food-rich conditions are followed by dryness and scarcity.
Biology is replete with life histories of comparable or even greater complexity. In protist life histories—by far the most diverse, exotic, and unique—one can search for ancestral modes of life, including missing links between the prokaryotes and eukaryotes. The one-celled swimming stage is called a sperm, whose imperative it is to find another one-celled partner, the ovum. Like all animals, humans develop from a single fertilized egg with its complement of two sets of genetic material. These diploid fertilized egg cells then divide to form many presumably identical cells. The early embryology of all animals goes through stages that have 2, 4, 8, 16, and so on cells. Genetic information is theoretically identical in each cell. But then how does it ever happen that, as they mature, the cells become permanently specialized to form hair,bone,liver,blood, or nerve cells? How does any given cell “know” what sort of specialized cell it must become, since all cells seem to contain identical nucleic acids? Despite a century of work on this process (called differentiation) and the discovery of many facts about embryos, this basic problem still remains unsolved in animal biology.
Fertilization of a human egg. (1) The sperm release enzymes that help disperse the corona radiata and bind to the zona pellucida. (2) The outer sperm head layer is sloughed off, exposing enzymes that digest a path through the zona pellucida. (3) The sperm fuses with the egg cell membrane, causing the zona pellucida to become impenetrable to other sperm. (4) The tail separates from the sperm head, and the male pronucleus enlarges and travels to the female pronucleus in the center of the cell. Chromosomes merge to form a fertilized egg.
Prior to the recognition of microbial life, the living world was too easily divided into animals that moved in pursuit of food and plants that produced food from sunlight. The futility of this simplistic classification scheme has been underscored by entire fields of science. Many bacteria both swim (like animals) and photosynthesize (like plants), yet they are best considered neither. Many algae (e.g., euglenids, dinomastigotes, chlorophytes) also swim and photosynthesize simultaneously. Molecular biological measurements of the DNA that codes for components of the ribosomes (organelles that are universally distributed) consistently find fungi to be extremely different from plants. Indeed, fungi genetically resemble animals more than plants.
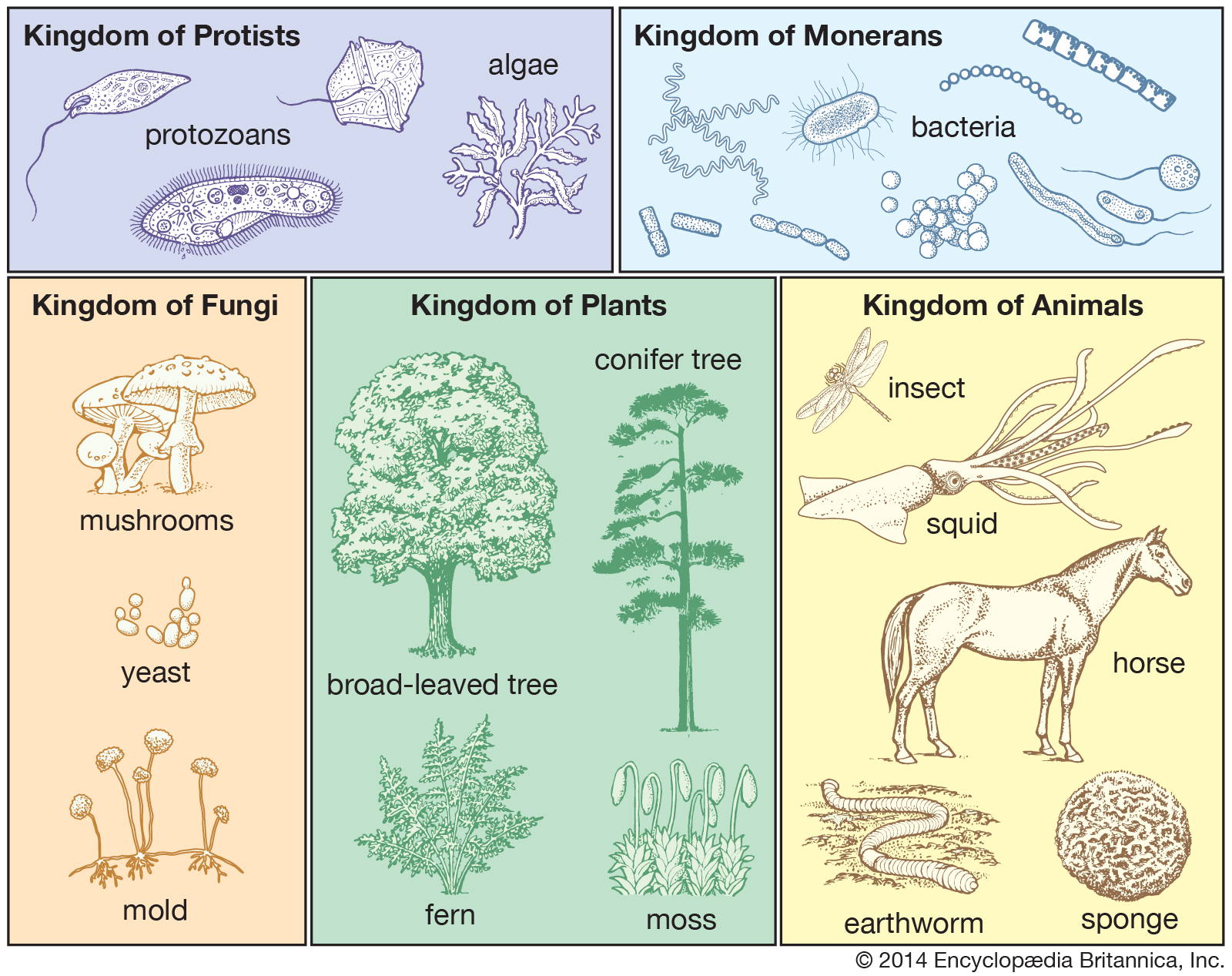
The Whittaker five-kingdom classification of life.
Modern biology, following the lead of the German biologist Ernst Haeckel and the American biologists Herbert F. Copeland and Robert H. Whittaker, has now thoroughly abandoned the two-kingdom plant-versus-animal dichotomy. Haeckel proposed three kingdoms when he established “Protista” for microorganisms. Copeland classified the microorganisms into the Monerans (prokaryotes) and the Protoctista (which included fungi with the rest of the eukaryotic microorganisms). His four-kingdom scheme (Monera, Protoctista, Animalia, and Plantae) had the advantage of clearly separating microbes with nuclei (Protoctista) from those without (Monera: the prokaryotes—that is, the bacteria and archaea) and of distinguishing the two embryo-forming groups—plants and animals—from the rest of life. Whittaker, on ecological grounds, raised the fungi to kingdom status. The modified Whittaker five-kingdom classification system is perhaps the most comprehensible and biologically based way to unambiguously organize information about all groups of living beings. American microbiologist Carl Woese has offered still another classification scheme, in which all organisms are placed in either the Archaea (prokaryotes that include some salt lovers, acid lovers, and methane producers), the Bacteria (all other prokaryotes, including obligate anaerobic bacteria as well as photosynthetic and chemoautotrophic bacteria), or the Eukarya (all eukaryote forms of life). Woese’s scheme is based on molecular biological criteria that focus on the RNA sequence of morphological factors to classify new or disputed organisms. Although Woese’s three-domain system is very popular, a potential problem with it is that RNA, one characteristic among thousands, does not consistently correlate with many others.
Microbes (or microbiota) are simply all those organisms too small to be visualized without some sort of microscopy. Bacteria, the smaller fungi, and the smaller protists are undoubtedly microbes. Some scientists classify tiny animals, worms, and rotifers as microbes as well. Like weed, a plant not wanted in a garden, microbe is often a more useful term than one with a precise scientific meaning.
The world of microbes, in any case, is more vast, complex, diverse, and widespread than the visible ordinary world of plants and animals. For example, microbes have sexual lives that are different from those of the animal and plant kingdoms. In all organisms composed of prokaryotic cells, DNA that is not complexed with protein (“naked,” or chromonemal, DNA) transfers from a source (such as a plasmid, a virus, a second cell, or even DNA molecules suspended in a solution) to a live prokaryotic cell. The recipient cell at the end of the sex act contains some quantity of its own DNA and integrates some from the donor. All prokaryotes can reproduce in the absence of any sex act.
In eukaryotes the sexual act requires the opening of membranes and the fusion of entire cells or at least of cell nuclei. The contribution from genes to the recombinant offspring is approximately 50 percent from each parent. From two to a dozen or so genders (in some species of paramecia) are present in any given sexual group. Although any given sex act requires at least two individuals, mating tends to be by pairs. Gender is understood to be those traits that predispose any organism to enter the sex act with any other. In multigender species only two genders or mating types enter a sex act at any one time. The rule is that in multigender species a mating requires any gender other than one’s own. Individual cells or multicellular organisms of complementary genders, in principle, produce fertile offspring. The universal rule is that no offspring result from matings of individuals of the same gender. In protists and fungi, uniparental reproduction (i.e., reproduction of a single parent) can occur in the absence of any sexual act, but two-parent sex may prevail seasonally or under other given environmental conditions in many inclusive taxa (such as families, classes, or phyla). Members of all species of the plant and animal kingdoms develop from embryos that form from a sexual act between the parents, and therefore two-parent (biparental) sex is the rule. Biparental sexuality of plants and animals has likely preceded its loss in all cases where a plant or animal species has reverted to uniparental reproduction, as in rotifers, whiptail lizards, and hundreds of plants that reproduce by runners rather than by seed. This suggestion is based on the fact that, at the cell level, aspects of meiosis (required for two-parent sex) continue to occur..
Ploidy, the concept of the number of complete sets of genes organized into chromosomes, is inapplicable to prokaryotes. Ploidy in protists, depending on species, varies so greatly and regularly that it is obvious that sexual cycles evolved in this diverse group of eukaryotes. Fungal cell nuclei are haploid (one set of chromosomes) or dikaryotic (two distinct nuclei from two different parents, each with one set of chromosomes sharing the same cell). Plant cell nuclei have two sets of chromosomes (diploid, in the sporophyte generation) or one (haploid, in the gametophyte generation). Animal cell nuclei, except in the gametes (sperm and egg), tend to have two sets of chromosomes (they are diploid).
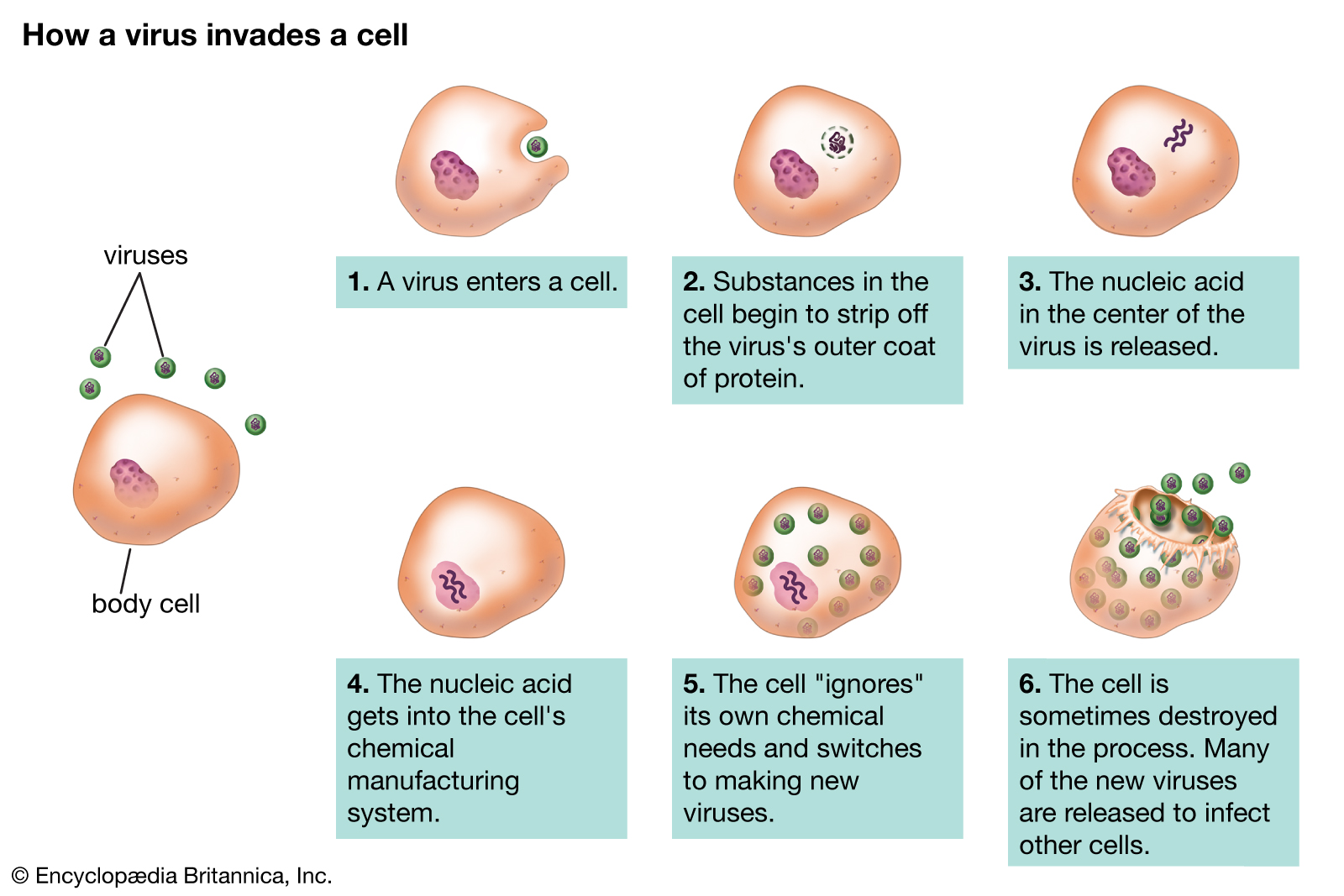
Viral and plasmidnucleic acids pass from cell to cell where the DNA or RNA perform their replication and coding functions efficiently. These “small replicons”—viruses and plasmids—are essentially strands of nucleic acid with a protein coat that depend entirely on the host cell for their continual existence. Pieces of the genetic material, virus-sized, pass from one cell into another cell of the same kind. Traveling small replicons of DNA produce genetic and permanently heritable changes in their new locations. Alternatively, part of the virus nucleic acid may be permanently bound to the nuclear DNA of the cell in which it resides. Viruses may be thought of as degenerate forms that are highly specialized in order to live in specific host cells of free-living organisms. Only cells are capable of performing the metabolic tasks that viruses and plasmids require. Viruses and plasmids must use the genetic transcription apparatus of cells. Bacterial viruses, or bacteriophages, may be extremely efficient in turning the luckless bacterium from a “factory” for the production of more bacteria into a factory for making more virus particles. It may take no more than 10 minutes for a bacterium infected by a single virus to produce 100 new virus particles, bursting forth from the victim bacterium by destroying it. Plasmids do not burst forth; rather, they benignly incorporate their DNA into that of their host cell.
Diagram showing a virus invading a cell and reproducing.
Organisms generated by the same fundamental biochemistry survive, grow, and reproduce in an extraordinarily wide range of conditions on Earth. For example, an alga called Cyanidium caldarium, a eukaryotic and photosynthetic organism, thrives in concentrated solutions of hot sulfuric acid and colours a damp landscape turquoise after a wet volcanic explosion. A swimming relative, Cyanophora paradoxa, survives in nearly these extremes. Certain less-colourful bacteria and fungi can live in extremely acidic environments (pH 0–2.5), such as that of Rio Tinto near Huelva in Spain. Bright blue-green cyanobacteria of many kinds can grow vigorously in extremely alkaline environments (pH 10–13).
Most familiar organisms on Earth are of course sensitive to extreme temperature in their surroundings. Mammals and birds have evolved internal regulation of their temperatures. Humans cannot tolerate body temperatures below 30 °C (86 °F) or above 40 °C (104 °F). Cold-climate organisms have special insulating layers of fat and fur. Other organisms adjust to seasonal temperature drops by developing dormant propagules such as spores, eggs, or tuns, which are hardy desiccation- and radiation-resistant forms produced by microscopic animals called tardigrades, also known as “water bears.” Dormancy is often accompanied by dehydration.
Most organisms are composed of an estimated 70–80 percent water. The availability of body water is a biological imperative. Certain halophilic bacteria live on water adsorbed on a single crystal of salt. Others such as the kangaroo rat (a mammal) and Tribolium (the flour beetle) imbibe no water at all in the liquid state. They rely entirely on metabolic water—that is, on water released from chemical bonds through the metabolism of food. A variety of plants, including Spanish moss, live without contact with groundwater. They extract water directly from the air, although they do require relatively high humidity. Desert plants and other plants in very dry environments, such as the two-leaved Welwitschia of the Namib Desert, have evolved extensive root systems that absorb subsurface water from a great volume of adjacent soil.
Life has been detected in the stratosphere and in the ocean’s major depths. Mud-loving photosynthetic bacteria live in pools at Yellowstone National Park at temperatures above 90 °C (194 °F), whereas some unidentified dark-dwelling marine bacteria at marine hot vents have been recorded to grow at a superhot temperature of 113 °C (235 °F). (Because the water is under pressure, it is not above the boiling point.) Sulfate-reducing bacteria taken from the ocean grow and reproduce at 104 °C (219 °F) under high hydrostatic pressure. Many organisms employ organic or inorganic antifreezes to lower the freezing point of their internal liquids. Many kinds can live at several tens of degrees below 0 °C (32 °F). Some insects use dimethylsulfoxide as an antifreeze. Other organisms live in briny pools in which dissolved salts lower the freezing point. San Juan Pond in Antarctica, for example, contains about one molecule of calcium chloride for every two water molecules. Not until –45 °C (–49 °F) does the pond freeze. A type of cryophilic (cold-loving) bacteria that lives there continues to metabolize down to at least –23 °C (–9 °F). Biological activity does not cease at the freezing point of water. In certain sea urchins, some microtubule proteins form the tubules of mitosis best at –2 °C (28 °F), and some enzymes are actually more active in ice than in water. Many bacteria are routinely frozen at –80 °C (–112 °F). They are thawed with no decrease in activity. Freezing temperatures alone cause no damage. Rather, frozen water removes tissue fluidity and leaves dangerous salt concentrations in its wake. The combination of expansion and contraction attendant to freezing and thawing harms membranes. Some arthropods can be severely dehydrated and then revived simply by the addition of water. Once dehydrated, these animals can be brought to any temperature from close to absolute zero (–273 °C, or –460 °F) to above the boiling point of water (100 °C, or 212 °F) without apparent damage. When encysted in response to dehydration, these arthropods at first glance are indistinguishable from a weathered grain of sand.
Cyanobacteria in a hot spring at Yellowstone National Park, Wyoming
Bacteria and fungal spores have been discovered near the base of the stratosphere by balloon searches. Organisms sought at much higher altitudes (up to 30,000 metres [100,000 feet]) have been detected; they are few in number and are all propagules. Birds have been observed to fly at maximum altitudes of 8,200 metres (27,000 feet), and on Mount Everest jumping spiders have been found at 6,700 metres (22,000 feet). At the opposite extreme, ciliates, pout fish, crabs, and clams have been recovered from ocean depths where pressures are hundreds of times those found at sea level. At these depths no light penetrates, and the organisms, some of which are quite large with bioluminescent organs that glow in the dark, feed on particles of organic matter raining down from the upper reaches of the oceans. Others sustain themselves by their chemoautotrophic bacterial associations.
The radiation environment of Earth has provoked evolutionary responses in many types of organisms. Some bacteria are readily killed by the small amount of solar ultraviolet light that filters through Earth’s atmosphere at wavelengths near 300 nanometres. To the continuing annoyance of nuclear physicists, the bacterium Deinococcus radiodurans thrives in the cooling pool of nuclear reactors amid radioactivity levels lethal to mammals. Some life avoids radiation by shielding: algae and some desert plants live under a superficial coating of soil or rock that is more transparent to visible light than to ultraviolet light. Many produce protective epithelial coatings. Most telling is the fact that some microbes and animals have active methods of repairing damage produced by radiation. Some of these repair mechanisms work in the dark; others require visible light. Nucleic acids of all organisms absorb ultraviolet light very effectively at a wavelength near 260 nanometres, which accounts for their ultraviolet sensitivity. The upper limit to the amount of ionizing radiation (which includes gamma rays,X-rays, and electrons) that an organism can receive without being killed is approximately 1,000,000 roentgens. Such an extraordinarily high dose can be withstood only by Deinococcus. Mammals are killed by vastly lower doses, probably because so much more can go wrong in a large and complex animal. For the whole body of a human being, a dose of some 400 roentgens causes radiation sickness and death in half of those exposed to this level. A thermonuclear weapon dropped on a populated area may deliver, through direct radiation and fallout, doses of a few hundred roentgens or more to people within a radius of some tens of kilometres of the target. Much smaller doses produce a variety of diseases as well as deleterious mutations in the hereditary material, the DNA of the chromosomes. The effect of small doses of radiation is apparently cumulative. Until very recently no human beings had lived in environments with large fluxes of ionizing radiation (seeradiation: Biologic effects of ionizing radiation).
A survey of the largest known single living organisms on Earth, which includes representatives from the animal, plant, and fungal kingdoms.
General Grant tree
General Grant tree, a giant sequoia (Sequoiadendron giganteum), among the largest trees in total bulk.
The sizes of organisms on Earth vary greatly and are not always easy to estimate. On the large end, great stands of poplar trees entirely connected by common roots are really a single organism. A variety of influences place an upper limit to the size of organisms. One is the strength of biological materials. Sequoia redwood trees, some of which exceed 90 metres (300 feet), are apparently near the upper limit of height for an organism. The Italian astronomer Galileo calculated in 1638 that a tree taller than roughly 90 metres would buckle under its own weight when displaced slightly from the vertical (for example, by a breeze). Because of the buoyancy of water, large animals such as whales are not presented with such stability problems. Other size-related difficulties arise. The volume of tissues to be nourished increases as the cube of the characteristic length of the organism, but the surface of the gut, which absorbs the ingested food, increases only as the square of the length for a fixed morphology. As an organism’s length is increased, a point of diminishing returns is ultimately reached where nutrition is irreversibly impeded in an animal.
New work on genome sequences, the total amount and quality of all of the genes that make up a live being, permits more accurate assessment of the material basis of the theoretically smallest and simplest extant free-living organisms. The complete DNA sequences of a few extremely small free-living organisms are now known—e.g., Mycoplasma genitalium with its 480 genes. All the molecules necessary for metabolism must be present. The smallest free-living cells include the pleuropneumonia-like organisms (PPLOs). Whereas an amoeba has a mass of 5 × 10−7 gram (2 × 10−8 ounce), a PPLO, which cannot be seen without a high-powered electron microscope, weighs 5 × 10−16 gram (2 × 10−15 ounce) and is only about 100 nanometres across. PPLOs grow very slowly. Other, even smaller organisms that grow even more slowly would be extremely difficult to detect. An organism the size of a PPLO that has room for only about a hundred enzymes depends entirely upon the animal tissue in which it lives. A much smaller organism would have room for many fewer enzymes. Its ability to accomplish the functions required for autopoiesis in living systems would be severely compromised. Were there, however, an environment in which all the necessary organic building blocks and such energy sources as ATP were provided “free,” then there might be a functioning organism substantially smaller than a PPLO. The inside of cells provides just such an environment, which explains why infectious agents, such as prions,plasmids, and viruses, may be substantially smaller than PPLOs. But it must be emphasized that viruses and their kin are not, even in principle, autopoietic.
The range of organic molecules that organisms, especially microbes, can metabolize is very wide and occasionally includes foods such as formaldehyde or petroleum that seem unlikely from a human point of view. Pseudomonas bacteria are capable of using almost any organic molecule as a source of carbon and energy, provided only that the molecule is at least slightly soluble in water. Microorganisms cannot metabolize plastics, not because of any fundamental chemical prohibitions but probably because plastics have not been part of the environment of microorganisms for very long. A lack of oxygen is thought of as extremely deleterious to life, but this view is anthropocentric. Many bacteria are facultative anaerobes that can take their oxygen or leave it. Many other bacteria and protists are obligate anaerobes that are actually poisoned by oxygen.
Water, which is crucial for life, is the major molecule in all organisms. Unless a massive mineral skeleton is present, the dry matter of most organisms is about one-half carbon by weight. This reflects the fact that all organic molecules are composed of carbon bound at least to hydrogen. Metabolism uses a wide variety of other chemical elements. Amino acids are made of nitrogen and sulfur in addition to carbon, hydrogen, and oxygen. Nucleic acids are made of phosphorus in addition to hydrogen, nitrogen, oxygen, and carbon. Sodium,potassium, and calcium are used to maintain electrolyte balance and to signal cells. Silicon is used as a structural material in the diatom shell, the radiolarian and heliozoanspicule, and the chrysophyte exoskeleton.Iron plays a fundamental role in the transport of molecular oxygen as part of the hemoglobin molecule. In some ascidians (sea squirts), however, vanadium replaces iron. Ascidian blood also contains unusually large amounts of niobium,titanium,chromium,manganese,molybdenum, and tungsten. The vanadium and niobium compounds in ascidian blood may be adaptations to low oxygen levels. Some bacteria use selenium,tellurium, or even arsenic as electron acceptors. Others produce the fully saturated gas hydrides of carbon, arsenic, phosphorus, or silicon as a metabolic waste. Still others form compounds of carbon with such halogens as chlorine or iodine. Not only the foregoing elements but also copper,zinc,cobalt, and possibly gallium,boron, and scandium perform particular functions in the enzymatic apparatus of particular cells. These elements, both the uncommon ones and those as common as phosphorus, are much more concentrated in living matter than in the environment where the living matter resides. This concentration suggests that such rare chemicals play unique functional roles that other, more abundant elements cannot serve.
Although any given organism is severely limited in its range of behaviour patterns and sensory capabilities, life as a whole is remarkably sensitive to aspects of its local social and physical environment. A bird raised from the egg in the absence of other members of its species migrates when the season beckons, builds the proper nest, and engages in elaborate courtship rituals. Those birds that fail to perpetuate the behaviour pattern do not leave descendants. Such behavioural accuracy itself must have evolved. Rats that pass through mazes easily interbreed, as do rats that pass through with difficulty; eventually two populations with inherited characteristics called “maze-smart” and “maze-dumb” are produced. Fruit fly populations attracted to light can be separated from those that avoid light. Classical genetic-crossing experiments reveal that the two populations differ largely in a small number of genes for phototropism. Similar genetic determinants of behaviour exist in humans. For example, possession of a supernumerary Y-chromosome in males is strikingly correlated with aggressive tendencies.
Arctic tern (Sterna paradisaea). It breeds in the southerly reaches of the Arctic and winters in the Antarctic regions, thus making the longest annual migration of any bird.
Photosensitivity, audiosensitivity, thermosensitivity, chemosensitivity, and magnetosensitivity
Photosensitivity, audiosensitivity, thermosensitivity, chemosensitivity, and magnetosensitivity (B)
An evening primrose (Oenothera biennis) seen (top) in visible light and (bottom) in ultraviolet light; the latter reveals nectar-guide patterns that are discernible to the moth pollinating this flower but not to the human eye.
Epauletted fruit bat (Epomophorus wahlbergi).
Silkworm moths (Bombyx mori) mating on cocoons.
Humans use only a limited region of the electromagnetic spectrum, the part called visible light, which extends from 400 to 700 nanometres in wavelength. While plants, algae, photosynthetic bacteria, and most animals are sensitive to this same range of wavelengths, many are sensitive to other wavelengths as well. Many plants present flower patterns visible only in the ultraviolet range at wavelengths below 400 nanometres, where pollinating insects are sensitive. Honeybees use polarized light—which the unaided human eye is unable to detect—for direction finding on partly cloudy days. The “pit” of such pit vipers as the rattlesnake is an infrared (heat) receptor that serves as a direction finder. These reptiles sense the thermal radiation emitted by mammals and birds, their warm-blooded prey. Humans are entirely insensitive to this thermal radiation.
That some animals such as dogs are sensitive to sounds that the human ear cannot detect is obvious to those who use dog whistles. Bats emit and detect sound waves at ultrahigh frequencies, in the vicinity of 100,000 cycles per second, about five times the highest frequency to which the human ear is sensitive. Bats have echolocated their prey by use of these sounds for millions of years before humans invented radar and sonar. The audio receptors of many moths that are prey to bats respond only to the frequencies emitted by the bats. When the bat sounds are heard, the moths take evasive action. Dolphins communicate via a very wide frequency range. They employ a “click” echolocator.
Some species of animals enjoy highly specialized and exotic organs for the detection or transmission of sound. Dolphins and whales use their blowholes rather than their mouths to utter their sounds.
Smell and taste, or some form of detection of specific chemical molecules, are universal. The ultimate in olfactory specialization may be male moths, whose feathery antennae are underlain by splayed microtubules, each of which is covered by a membrane at the distal end. They smell essentially nothing except the epoxide compound called disparlure, the chemical sex attractant discharged by the female. Only 40 molecules per second need impact on the antennae to produce a marked response. One female silkworm moth need release only 10–8 gram (4 × 10–10 ounce) of sex attractant per second in order to attract every male silkworm moth within a few kilometres.
Magnetotactic bacteria sense Earth’s magnetic field. North Pole-seeking bacteria swim toward the sediment-water interface as they follow the magnetic lines of force. South Pole-seeking flagellated magnetotactic bacteria do the same in the Southern Hemisphere. Since those studied are microaerophiles—i.e., they require oxygen in lower than ambient concentrations—pole seekers tend to arrive at oxygen-depleted sediment adequate for their continued growth and reproduction. Ultrastructural studies reveal magnetosomes, tiny single-domain crystals of magnetite, an iron oxide mineral sensitive to magnetic fields, or greigite, an iron sulfide mineral, in their cells. The magnetosomes are aligned along the axis of the cell and serve to orient the sensitive bacteria. All the different kinds of magnetotactic bacteria bear magnetosomes in their cells. Whether magnetotaxis is causal in the orientation of homing pigeons, dancing bees on cloudy days, or other instinctively orienting animals is under investigation.
Besides the familiar senses of sight, hearing, smell, taste, and touch, organisms have a wide variety of other senses (see aboveSensory capabilities and awareness). People have inertial orientation systems and accelerometers in the cochlear canal of the ear. The water scorpion (Nepa) has a fathometer sensitive to hydrostatic pressure gradients. Many plants have chemically amplified gravity sensors made of modified chloroplasts. Some green algae use barium sulfate and calcium ion detection systems to sense gravity. Fireflies and squids communicate with their own kind by producing changing patterns of light on their bodies. The nocturnal African freshwater fish Gymnarchus niloticus operates a dipole electrostatic field generator and a sensor to detect the amplitude and frequency of disturbances in turbulent waters.
From the foregoing sample of sensibilities, it is clear that a vast repertoire of sensors in living beings confers upon their possessors an awareness of the environment that differs from humanity’s. Humans, however, have an enhanced ability to extend their sensory and intellectual capabilities through the use of instrumentation far beyond those with which they are born.
The sensory system of Earth is expanded by the capabilities of human machines. From those that detect ionizing radiation, wind velocities, the taste of wine, the concentration of salt in solution, a few photons of light in a dark corridor, or the blood temperature of an infant to those that record microearthquakes, a lying smile, or the heat of a furnace, the sensory systems of the biosphere—nonhuman, human, and human-mediated—have augmented over time. Indeed, just as seeing-eye dogs transmit visual information to their blind owners, the sensory system of life extends far beyond any given species of animals and its machines to the entire sensitive biota in this pulsating biosphere. Sensitivity to sound, chemicals, heat, light, mechanical movement, magnetism, and charged particles has been tallied by many a hardworking scientist. Whether entire categories of sensory information are missing from that list is not entirely clear. Great sensitivity to the environment abounds even in those smallest life-forms, the bacteria. Life has been sensing and responding to its environment since its inception more than 3.7 billion to 3.5 billion years ago. Moreover, it is not clear at what point in evolutionary history, or where precisely among organisms, consciousness comes in. Humans are conscious and self-conscious. But are protists that choose certain shapes and sizes of glass beads over others conscious of their decision making? Charles Darwin recognized selection among various male suitors by females as instrumental in the evolution of sexual species, including birds and insects. The extent to which consciousness and choice making are important in evolution remains a matter for debate.
Evolution and the history of life on Earth
Evolution and the history of life on Earth
Evolution and the history of life on Earth (B)
No text.
The evidence is overwhelming that all life on Earth has evolved from common ancestors in an unbroken chain since its origin. Darwin’s principle of evolution is summarized by the following facts. All life tends to increase: more organisms are conceived, born, hatched, germinated from seed, sprouted from spores, or produced by cell division (or other means) than can possibly survive. Each organism so produced varies, however little, in some measurable way from its relatives. In any given environment at any given time, those variants best suited to that environment will tend to leave more offspring than the others. Offspring resemble their ancestors. Variant organisms will leave offspring like themselves. Therefore, organisms will diverge from their ancestors with time. The term natural selection is shorthand for saying that all organisms do not survive to leave offspring with the same probability. Those alive today have been selected relative to similar ones that never survived or procreated. All organisms on Earth today are equally evolved since all share the same ancient original ancestors who faced myriad threats to their survival. All have persisted since roughly 3.7 billion to 3.5 billion years ago during the Archean Eon (4 billion to 2.5 billion years ago), products of the great evolutionary process with its identical molecular biological bases. Because the environment of Earth is so varied, the particular details of any organism’s evolutionary history differ from those of another species in spite of chemical similarities.
The geologic time scale from 650 million years ago to the present, showing major evolutionary events.
Everywhere the environment of Earth is heterogeneous. Mountains, oceans, and deserts suffer extremes of temperature, humidity, and water availability. All ecosystems contain diverse microenvironments: oxygen-depleted oceanic oozes, sulfide- or ammonia-rich soils, mineral outcrops with a high radioactivity content, or boiling organic-rich springs, for example. Besides these physical factors, the environment of any organism involves the other organisms in its surroundings. For each environmental condition, there is a corresponding ecological niche. The variety of ecological niches populated on Earth is quite remarkable. Even wet cracks in granite are replete with “rock eating” bacteria. Ecological niches in the history of life have been filled independently several times. For example, quite analogous to the ordinary placental mammalian wolf was the marsupial wolf, the thylacine (extinct since 1936) that lived in Australia; the two predatory mammals have striking similarities in physical appearance and behaviour. The same streamlined shape for high-speed marine motion evolved independently at least four times: in Stenopterygius and other Mesozoic reptiles; in tuna, which are fish; and in dolphins and seals, which are mammals. Convergent evolution in hydrodynamic form arises from the fact that only a narrow range of solutions to the problem of high-speed marine motion by large animals exists. The eye, a light receptor that makes an image, has evolved independently more than two dozen times not only in animals on Earth but in protists such as the dinomastigote Erythropsodinium. Apparently eyelike structures best solve the problem of visual recording. Where physics or chemistry establishes one most efficient solution to a given ecological problem, evolution in distinct lineages will often tend toward similar, nearly identical solutions. This phenomenon is known as convergent evolution.
Parallel evolution of marsupial and placental mammals
Parallel evolution of marsupial mammals in Australia and placental mammals on other continents.
Life ultimately is a material process that arose from a nonliving material system spontaneously—and at least once in the remote past. How life originated is discussed below. Yet no evidence for spontaneous generation now can be cited. Spontaneous generation, also called abiogenesis, the hypothetical process by which living organisms develop from nonliving matter, must be rejected. According to this theory, pieces of cheese and bread wrapped in rags and left in a dark corner were thought to produce mice, because after several weeks mice appeared in the rags. Many believed in spontaneous generation because it explained such occurrences as maggots swarming on decaying meat.
By the 18th century it had become obvious that plants and animals could not be produced by nonliving material. The origin of microorganisms such as yeast and bacteria, however, was not fully determined until French chemist Louis Pasteur proved in the 19th century that microorganisms reproduce, that all organisms come from preexisting organisms, and that all cells come from preexisting cells. Then what evidence is there for the earliest life on Earth?
Past time on Earth, as inferred from the rock record, is divided into four immense periods of time called eons. These are the Hadean (4.6 billion to 4 billion years ago), the Archean (4 billion to 2.5 billion years ago), the Proterozoic (2.5 billion to 541 million years ago), and the Phanerozoic (541 million years ago to the present). For the Hadean Eon, the only record comes from meteorites and lunar rocks. No rocks of Hadean age survive on Earth. In the figure, eons are divided into eras, periods, and epochs. Such entries in the geologic time scale are often called “geologic time intervals.”
Perhaps the most fundamental and at the same time the least understood biological problem is the origin of life. It is central to many scientific and philosophical problems and to any consideration of extraterrestrial life. Most of the hypotheses of the origin of life will fall into one of four categories:
The origin of life is a result of a supernatural event—that is, one irretrievably beyond the descriptive powers of physics,chemistry, and other science.
Life, particularly simple forms, spontaneously and readily arises from nonliving matter in short periods of time, today as in the past.
Life is coeternal with matter and has no beginning; life arrived on Earth at the time of Earth’s origin or shortly thereafter.
Life arose on the early Earth by a series of progressive chemical reactions. Such reactions may have been likely or may have required one or more highly improbable chemical events.
The Creation of Adam, detail of the ceiling fresco by Michelangelo, 1508–12; in the Sistine Chapel, Vatican City.
Hypothesis 1, the traditional contention of theology and some philosophy, is in its most general form not inconsistent with contemporary scientific knowledge, although scientific knowledge is inconsistent with a literal interpretation of the biblical accounts given in chapters 1 and 2 of Genesis and in other religious writings. Hypothesis 2 (not of course inconsistent with 1) was the prevailing opinion for centuries. A typical 17th-century view follows:
“[May one] doubt whether, in cheese and timber, worms are generated, or, if beetles and wasps, in cow’s dung, or if butterflies, locusts, shellfish, snails, eels, and suchlike be procreated of putrefied matter, which is apt to receive the form of that creature to which it is by the formative power disposed. To question this is to question reason, sense, and experience. If he doubts of this, let him go to Egypt, and there he will find the fields swarming with mice begot of the mud of the Nylus [Nile], to the great calamity of the inhabitants.”
(Alexander Ross, Arcana Microcosmi, 1652.)
It was not until the Renaissance, with its burgeoning interest in anatomy, that such spontaneous generation of animals from putrefying matter was deemed impossible. During the mid-17th century the British physiologist William Harvey, in the course of his studies on the reproduction and development of the king’s deer, discovered that every animal comes from an egg. An Italian biologist, Francesco Redi, established in the latter part of the 17th century that the maggots in meat came from flies’ eggs, deposited on the meat. In the 18th century an Italian priest, Lazzaro Spallanzani, showed that fertilization of eggs by sperm was necessary for the reproduction of mammals. Yet the idea of spontaneous generation died hard. Even though it was clear that large animals developed from fertile eggs, there was still hope that smaller beings, microorganisms, spontaneously generated from debris. Many felt it was obvious that the ubiquitous microscopic creatures generated continually from inorganic matter.
Maggots were prevented from developing on meat by covering it with a flyproof screen. Yet grape juice could not be kept from fermenting by putting over it any netting whatever. Spontaneous generation was the subject of a great controversy between the famous French bacteriologists Louis Pasteur and Félix-Archimède Pouchet in the 1850s. Pasteur triumphantly showed that even the most minute creatures came from “germs” that floated downward in the air, but that they could be impeded from access to foodstuffs by suitable filtration. Pouchet argued, defensibly, that life must somehow arise from nonliving matter; if not, how had life come about in the first place?
Pasteur’s experimental results were definitive: life does not spontaneously appear from nonliving matter. American historian James Strick reviewed the controversies of the late 19th century between evolutionists who supported the idea of “life from non-life” and their responses to Pasteur’s religious view that only the Deity can make life. The microbiological certainty that life always comes from preexisting life in the form of cells inhibited many post-Pasteur scientists from discussions of the origin of life at all. Many were, and still are, reluctant to offend religious sentiment by probing this provocative subject. But the legitimate issues of life’s origin and its relation to religious and scientific thought raised by Strick and other authors, such as the Australian Reg Morrison, persist today and will continue to engender debate.
Toward the end of the 19th century, hypothesis 3 gained currency. Swedish chemist Svante A. Arrhenius suggested that life on Earth arose from “panspermia,” microscopic spores that wafted through space from planet to planet or solar system to solar system by radiation pressure. This idea, of course, avoids rather than solves the problem of the origin of life. It seems extremely unlikely that any live organism could be transported to Earth over interplanetary or, worse yet, interstellar distances without being killed by the combined effects of cold, desiccation in a vacuum, and radiation.
Although English naturalist Charles Darwin did not commit himself on the origin of life, others subscribed to hypothesis 4 more resolutely. The famous British biologist T.H. Huxley in his book Protoplasm: The Physical Basis of Life (1869) and the British physicist John Tyndall in his “Belfast Address” of 1874 both asserted that life could be generated from inorganic chemicals. However, they had extremely vague ideas about how this might be accomplished. The very phrase “organic molecule” implied, especially then, a class of chemicals uniquely of biological origin. Despite the fact that urea and other organic (carbon-hydrogen)molecules had been routinely produced from inorganic chemicals since 1828, the term organic meant “from life” to many scientists and still does. In the following discussion the word organic implies no necessary biological origin. The origin-of-life problem largely reduces to determination of an organic, nonbiological source of certain processes such as the identity maintained by metabolism, growth, and reproduction (i.e., autopoiesis).
Darwin’s attitude was: “It is mere rubbish thinking at present of the origin of life; one might as well think of the origin of matter.” The two problems are in fact curiously connected. Indeed, modern astrophysicists do think about the origin of matter. The evidence is convincing that thermonuclear reactions, either in stellar interiors or in supernova explosions, generate all the chemical elements of the periodic table more massive than hydrogen and helium. Supernova explosions and stellar winds then distribute the elements into the interstellar medium, from which subsequent generations of stars and planets form. These thermonuclear processes are frequent and well-documented. Some thermonuclear reactions are more probable than others. These facts lead to the idea that a certain cosmic distribution of the major elements occurs throughout the universe. Some atoms of biological interest, their relative numerical abundances in the universe as a whole, on Earth, and in living organisms are listed in the table. Even though elemental composition varies from star to star, from place to place on Earth, and from organism to organism, these comparisons are instructive: the composition of life is intermediate between the average composition of the universe and the average composition of Earth. Ninety-nine percent of the mass both of the universe and of life is made of six atoms: hydrogen (H), helium (He), carbon (C), nitrogen (N), oxygen (O), and neon (Ne). Might not life on Earth have arisen when Earth’s chemical composition was closer to the average cosmic composition and before subsequent events changed Earth’s gross chemical composition?
*0 percent here stands for any quantity less than 10–6 percent.
Photograph of Jupiter taken by Voyager 1 on February 1, 1979, at a range of 32.7 million km (20.3 million miles). Prominent are the planet's pastel-shaded cloud bands and Great Red Spot (lower centre).
The Jovian planets (Jupiter,Saturn,Uranus, and Neptune) are much closer to cosmic composition than is Earth. They are largely gaseous, with atmospheres composed principally of hydrogen and helium. Methane,ammonia, neon, and water have been detected in smaller quantities. This circumstance very strongly suggests that the massive Jovian planets formed from material of typical cosmic composition. Because they are so far from the Sun, their upper atmospheres are very cold. Atoms in the upper atmospheres of the massive, cold Jovian planets cannot now escape from their gravitational fields, and escape was probably difficult even during planetary formation.
Earth and the other planets of the inner solar system, however, are much less massive, and most have hotter upper atmospheres. Hydrogen and helium escape from Earth today; it may well have been possible for much heavier gases to have escaped during Earth’s formation. Very early in Earth’s history, there was a much larger abundance of hydrogen, which has subsequently been lost to space. Most likely the atoms carbon, nitrogen, and oxygen were present on the early Earth, not in the forms of CO2 (carbon dioxide), N2, and O2 as they are today but rather as their fully saturated hydrides: methane, ammonia, and water. The presence of large quantities of reduced (hydrogen-rich) minerals, such as uraninite and pyrite, that were exposed to the ancient atmosphere in sediments formed over two billion years ago implies that atmospheric conditions then were considerably less oxidizing than they are today.
In the 1920s British geneticist J.B.S. Haldane and Russian biochemist Aleksandr Oparin recognized that the nonbiological production of organic molecules in the present oxygen-rich atmosphere of Earth is highly unlikely but that, if Earth once had more hydrogen-rich conditions, the abiogenic production of organic molecules would have been much more likely. If large quantities of organic matter were somehow synthesized on early Earth, they would not necessarily have left much of a trace today. In the present atmosphere—with 21 percent of oxygen produced by cyanobacterial, algal, and plant photosynthesis—organic molecules would tend, over geological time, to be broken down and oxidized to carbon dioxide, nitrogen, and water. As Darwin recognized, the earliest organisms would have tended to consume any organic matter spontaneously produced prior to the origin of life.
The first experimental simulation of early Earth conditions was carried out in 1953 by a graduate student, Stanley L. Miller, under the guidance of his professor at the University of Chicago, chemist Harold C. Urey. A mixture of methane, ammonia, water vapour, and hydrogen was circulated through a liquid solution and continuously sparked by a corona discharge mounted higher in the apparatus. The discharge was thought to represent lightning flashes. After several days of exposure to sparking, the solution changed colour. Several amino and hydroxy acids, familiar chemicals in contemporary Earth life, were produced by this simple procedure. The experiment is simple enough that the amino acids can readily be detected by paper chromatography by high school students. Ultraviolet light or heat was substituted as an energy source in subsequent experiments. The initial abundances of gases were altered. In many other experiments like this, amino acids were formed in large quantities. On the early Earth much more energy was available in ultraviolet light than from lightning discharges. At long ultraviolet wavelengths, methane, ammonia, water, and hydrogen are all transparent, and much of the solar ultraviolet energy lies in this region of the spectrum. The gas hydrogen sulfide was suggested to be a likely compound relevant to ultraviolet absorption in Earth’s early atmosphere. Amino acids were also produced by long-wavelength ultraviolet irradiation of a mixture of methane, ammonia, water, and hydrogen sulfide. At least some of these amino acid syntheses involved hydrogen cyanide and aldehydes (e.g., formaldehyde) as gaseous intermediates formed from the initial gases. That amino acids, particularly biologically abundant amino acids, are made readily under simulated early Earth conditions is quite remarkable. If oxygen is permitted in these kinds of experiments, no amino acids are formed. This has led to a consensus that hydrogen-rich (or at least oxygen-poor) conditions were necessary for natural organic syntheses prior to the appearance of life.
Under alkaline conditions, and in the presence of inorganic catalysts, formaldehyde spontaneously reacts to form a variety of sugars. The five-carbon sugars fundamental to the formation of nucleic acids, as well as six-carbon sugars such as glucose and fructose, are easily produced. These are common metabolites and structural building blocks in life today. Furthermore, the nucleotide bases and even the biological pigments called porphyrins have been produced in the laboratory under simulated early Earth conditions. Both the details of the experimental synthetic pathways and the question of stability of the small organic molecules produced are vigorously debated. Nevertheless, most, if not all, of the essential building blocks of proteins (amino acids), carbohydrates (sugars), and nucleic acids (nucleotide bases)—that is, the monomers—can be readily produced under conditions thought to have prevailed on Earth in the Archean Eon. The search for the first steps in the origin of life has been transformed from a religious/philosophical exercise to an experimental science.
The formation of polymers, long-chain molecules made of repeating units of monomers (the essential building blocks mentioned above), is a far more difficult experimental problem than the formation of monomers. Polymerization reactions tend to be dehydrations. A molecule of water is lost in the formation of a peptide from two amino acids or of a disaccharid sugar from two monomers. Dehydrating agents are used to initiate polymerization. The polymerization of amino acids to form long proteinlike molecules (“proteinoids”) was accomplished through dry heating by American biochemist Sidney Fox and his colleagues. The polyamino acids that he formed are not random molecules unrelated to life. They have distinct catalytic activities. Long polymers of amino acids were also produced from hydrogen cyanide and anhydrous liquid ammonia by American chemist Clifford Matthews in simulations of the early upper atmosphere. Some evidence exists that ultraviolet irradiation induces combinations of nucleotide bases and sugars in the presence of phosphates or cyanides. Some condensing agents such as cyanamide are efficiently made under simulated primitive conditions. Despite the breakdown by water of molecular intermediates, condensing agents may quite effectively induce polymerization, and polymers of amino acids, sugars, and nucleotides have all been made this way.
That adsorption of relevant small carbon compounds on clays or other minerals may have concentrated these intermediates was suggested by the British scientist John Desmond Bernal. Concentration of some kind is required to offset the tendency for water to break down polymers of biological significance. Phosphorus, which with deoxyribose sugar forms the “backbone” of DNA and is integrally involved in cell energy transformation and membrane formation, is preferentially incorporated into prebiological organic molecules. It is hard to explain how such a preference could have happened without the concentration of organic molecules.
The early ocean and lakes themselves may have been a dilute solution of organic molecules. If all the surface carbon on Earth were present as organic molecules, or if many known ultraviolet synthetic reactions that produce organic molecules were permitted to continue for a billion years with their products dissolved in the oceans, a 1 percent solution of organic molecules would result. Haldane suggested that the origin of life occurred in a “hot dilute soup.” Concentration through either evaporation or freezing of pools, adsorption on clay interfaces, or the generation of colloidal enclosures called coacervates may have served to bring the organic molecules in question in contact with each other.
The essential building blocks for life (the monomers) were probably produced in relatively abundant concentrations, given conditions on the early Earth. Although relevant, this is more akin to the origin of food than to the origin of life. If life is defined as a self-maintaining, self-producing, and mutable molecular system that derives energy and supplies from the environment, then food is certainly required for life. Polynucleotides (polymers of RNA and DNA) can be produced in laboratory experiments from nucleotide phosphates in the presence of enzymes of biological origin (polymerases) and a preexisting “primer” nucleic acid molecule. If the primer is absent, polynucleotides are still formed, but they lack specific genetic information. Once such a polynucleotide forms, it can act as a primer for subsequent syntheses.
Even if such a molecular population could replicate polynucleotides, it would not be considered alive. The polynucleotides tend to hydrolyze (break down) in water. In the early 1980s American biochemist Thomas Cech and Canadian American molecular biologist Sidney Altman discovered that certain RNA molecules have catalytic properties. They catalyze their own splicing, which suggests an early role for RNA in life or even in life’s origins. Only the partnership of the two kinds of molecules (proteins and nucleic acids) segregated from the rest of the world by an oily membrane makes the growth process of life on Earth possible. The molecular apparatus ancillary to the operation of the genetic code—the rules that determine the linear order of amino acids in proteins from nucleotide base pairs in nucleic acids (i.e., the activating enzymes,transfer RNAs,messenger RNAs,ribosomes, and so on)—may be the product of a long evolutionary history among natural, thermodynamically favoured, gradient-reducing complex systems. These rules are produced according to instructions contained within the code. American biophysicist Harold J. Morowitz argued cogently that the origin of the genetic system, the code with its elaborate molecular apparatus, occurred inside cells only after the origin of life as a cyclic metabolic system. American theoretical biologist Jeffrey Wicken pointed out that replicating molecules, if they appeared first, would have had no impetus to develop a complex cellular package or associated protein machinery and that life thus probably arose as a metabolic system that was stabilized by the genetic code, which allowed life’s second law-favoured process to continue ad infinitum.
Many separate and rather diverse instances of the origin of living cells may have occurred in the Archean Earth, but obviously only one prevailed. Interactions eventually eliminated all but our lineage. From the common composition, metabolism, chemical behaviour, and other properties of life, it seems clear that every organism on Earth today is a member of the same lineage.
Most organic molecules made by living systems inside cells display the same optical activity: when exposed to a beam of plane-polarized light, they rotate the plane of the beam. Amino acids rotate light to the left, whereas sugars, called dextrorotatory, rotate it to the right. Organic molecules produced artificially lack optical activity because both “left-handed” and “right-handed” molecules are present in equal quantity. Molecules of the same optical activity can be assembled in complementary ways like the stacking of right-handed gloves. The same monomers can be used to produce longer chain molecules that are three-dimensional mirror images of each other; mixtures of monomers of different handedness cannot. Cumulative symmetry is responsible for optical activity. At the time of the origin of life, organic molecules, corresponding both to left- and right-handed forms, were no doubt formed as they are in laboratory simulation experiments today: both types were produced. But the first living systems must have employed one type of component, for the same reason that carpenters cannot use random mixtures of screws with left- and right-handed threads in the same project with the same tools. Whether left- or right-handed activity was adopted was probably a matter of chance, but, once a particular asymmetry was established, it maintained itself. Optical activity accordingly is likely to be a feature of life on any planet. The chances may be equal of finding a given organic molecule or its mirror image in extraterrestrial life-forms if, as Morowitz suspects, the incorporation of nitrogen into the first living system involved glutamine, the simplest of the required amino acid precursors with optical activity.
The first living cells probably resided in a molecular Garden of Eden, where the prebiological origin of food had guaranteed monomers that were available. The cells, the first single-celled organisms, would have increased rapidly. But such an increase was eventually limited by the supply of molecular building blocks. Those organisms with an ability to synthesize scarce monomers, say A, from more abundant ones, say B, would have persisted. The secondary source of supply, B, would in time also become depleted. Those organisms that could produce B from a third monomer, C, would have preferentially persisted. The American biochemist Norman H. Horowitz has proposed that the multienzyme catalyzed reaction chains of contemporary cells originally evolved in this way.
Abiogenesis is the natural process of life arising from non-living matter, such as simple organic compounds. The prevailing scientific hypothesis is that the transition from non-living to living entities was not a single event, but a gradual process of increasing complexity. Life on Earth first appeared as early as 4.28 billion years ago, soon after ocean formation 4.41 billion years ago, and not long after the formation of the Earth 4.54 billion years ago. The earliest known life forms are microfossils of bacteria. Researchers generally think that current life on Earth descends from an RNA world, although RNA-based life may not have been the first life to have existed. The classic 1952 Miller–Urey experiment and similar research demonstrated that most amino acids, the chemical constituents of the proteins used in all living organisms, can be synthesized from inorganic compounds under conditions intended to replicate those of the early Earth. Complex organic molecules occur in the Solar System and in interstellar space, and these molecules may have provided starting material for the development of life on Earth.
Since its primordial beginnings, life on Earth has changed its environment on a geologic time scale, but it has also adapted to survive in most ecosystems and conditions. Some microorganisms, called extremophiles, thrive in physically or geochemically extreme environments that are detrimental to most other life on Earth. The cell is considered the structural and functional unit of life. There are two kinds of cells, prokaryotic and eukaryotic, both of which consist of cytoplasm enclosed within a membrane and contain many biomolecules such as proteins and nucleic acids. Cells reproduce through a process of cell division, in which the parent cell divides into two or more daughter cells.
In the past, there have been many attempts to define what is meant by "life" through obsolete concepts such as odic force,hylomorphism,spontaneous generation and vitalism, that have now been disproved by biological discoveries.Aristotle is considered to be the first person to classify organisms. Later, Carl Linnaeus introduced his system of binomial nomenclature for the classification of species. Eventually new groups and categories of life were discovered, such as cells and microorganisms, forcing dramatic revisions of the structure of relationships between living organisms. Though currently only known on Earth, life need not be restricted to it, and many scientists speculate in the existence of extraterrestrial life.Artificial life is a computer simulation or human-made reconstruction of any aspect of life, which is often used to examine systems related to natural life.
Death is the permanent termination of all biological functions which sustain an organism, and as such, is the end of its life. Extinction is the term describing the dying out of a group or taxon, usually a species.Fossils are the preserved remains or traces of organisms.
The definition of life has long been a challenge for scientists and philosophers, with many varied definitions put forward. This is partially because life is a process, not a substance. This is complicated by a lack of knowledge of the characteristics of living entities, if any, that may have developed outside of Earth. Philosophical definitions of life have also been put forward, with similar difficulties on how to distinguish living things from the non-living. Legal definitions of life have also been described and debated, though these generally focus on the decision to declare a human dead, and the legal ramifications of this decision.
Since there is no unequivocal definition of life, most current definitions in biology are descriptive. Life is considered a characteristic of something that preserves, furthers or reinforces its existence in the given environment. This characteristic exhibits all or most of the following traits:
Homeostasis: regulation of the internal environment to maintain a constant state; for example, sweating to reduce temperature
Organization: being structurally composed of one or more cells – the basic units of life
Metabolism: transformation of energy by converting chemicals and energy into cellular components (anabolism) and decomposing organic matter (catabolism). Living things require energy to maintain internal organization (homeostasis) and to produce the other phenomena associated with life.
Growth: maintenance of a higher rate of anabolism than catabolism. A growing organism increases in size in all of its parts, rather than simply accumulating matter.
Adaptation: the ability to change over time in response to the environment. This ability is fundamental to the process of evolution and is determined by the organism's heredity, diet, and external factors.
Response to stimuli: a response can take many forms, from the contraction of a unicellular organism to external chemicals, to complex reactions involving all the senses of multicellular organisms. A response is often expressed by motion; for example, the leaves of a plant turning toward the sun (phototropism), and chemotaxis.
Reproduction: the ability to produce new individual organisms, either asexually from a single parent organism or sexually from two parent organisms.
From a physics perspective, living beings are thermodynamic systems with an organized molecular structure that can reproduce itself and evolve as survival dictates. Thermodynamically, life has been described as an open system which makes use of gradients in its surroundings to create imperfect copies of itself. Hence, life is a self-sustained chemical system capable of undergoing Darwinian evolution. A major strength of this definition is that it distinguishes life by the evolutionary process rather than its chemical composition.
Others take a systemic viewpoint that does not necessarily depend on molecular chemistry. One systemic definition of life is that living things are self-organizing and autopoietic (self-producing). Variations of this definition include Stuart Kauffman's definition as an autonomous agent or a multi-agent system capable of reproducing itself or themselves, and of completing at least one thermodynamic work cycle. This definition is extended by the apparition of novel functions over time.
Whether or not viruses should be considered as alive is controversial. They are most often considered as just replicators rather than forms of life. They have been described as "organisms at the edge of life" because they possess genes, evolve by natural selection, and replicate by creating multiple copies of themselves through self-assembly. However, viruses do not metabolize and they require a host cell to make new products. Virus self-assembly within host cells has implications for the study of the origin of life, as it may support the hypothesis that life could have started as self-assembling organic molecules.
To reflect the minimum phenomena required, other biological definitions of life have been proposed, with many of these being based upon chemical systems.Biophysicists have commented that living things function on negative entropy. In other words, living processes can be viewed as a delay of the spontaneous diffusion or dispersion of the internal energy of biological molecules towards more potential microstates. In more detail, according to physicists such as John Bernal,Erwin Schrödinger,Eugene Wigner, and John Avery, life is a member of the class of phenomena that are open or continuous systems able to decrease their internal entropy at the expense of substances or free energy taken in from the environment and subsequently rejected in a degraded form.
Living systems are open self-organizing living things that interact with their environment. These systems are maintained by flows of information, energy, and matter.
Budisa, Kubyshkin and Schmidt defined cellular life as an organizational unit resting on four pillars/cornerstones: (i) energy, (ii) metabolism, (iii) information and (iv) form. This system is able to regulate and control metabolism and energy supply and contains at least one subsystem that functions as an information carrier (genetic information).Cells as self-sustaining units are parts of different populations that are involved in the unidirectional and irreversible open-ended process known as evolution.
Some scientists have proposed in the last few decades that a general living systems theory is required to explain the nature of life. Such a general theory would arise out of the ecological and biological sciences and attempt to map general principles for how all living systems work. Instead of examining phenomena by attempting to break things down into components, a general living systems theory explores phenomena in terms of dynamic patterns of the relationships of organisms with their environment.
The idea that the Earth is alive is found in philosophy and religion, but the first scientific discussion of it was by the Scottish scientist James Hutton. In 1785, he stated that the Earth was a superorganism and that its proper study should be physiology. Hutton is considered the father of geology, but his idea of a living Earth was forgotten in the intense reductionism of the 19th century.:10 The Gaia hypothesis, proposed in the 1960s by scientist James Lovelock, suggests that life on Earth functions as a single organism that defines and maintains environmental conditions necessary for its survival. This hypothesis served as one of the foundations of the modern Earth system science.
The first attempt at a general living systems theory for explaining the nature of life was in 1978, by American biologist James Grier Miller.Robert Rosen (1991) built on this by defining a system component as "a unit of organization; a part with a function, i.e., a definite relation between part and whole." From this and other starting concepts, he developed a "relational theory of systems" that attempts to explain the special properties of life. Specifically, he identified the "nonfractionability of components in an organism" as the fundamental difference between living systems and "biological machines."
A systems view of life treats environmental fluxes and biological fluxes together as a "reciprocity of influence," and a reciprocal relation with environment is arguably as important for understanding life as it is for understanding ecosystems. As Harold J. Morowitz (1992) explains it, life is a property of an ecological system rather than a single organism or species. He argues that an ecosystemic definition of life is preferable to a strictly biochemical or physical one. Robert Ulanowicz (2009) highlights mutualism as the key to understand the systemic, order-generating behavior of life and ecosystems.
Complex systems biology (CSB) is a field of science that studies the emergence of complexity in functional organisms from the viewpoint of dynamic systems theory. The latter is also often called systems biology and aims to understand the most fundamental aspects of life. A closely related approach to CSB and systems biology called relational biology is concerned mainly with understanding life processes in terms of the most important relations, and categories of such relations among the essential functional components of organisms; for multicellular organisms, this has been defined as "categorical biology", or a model representation of organisms as a category theory of biological relations, as well as an algebraic topology of the functional organization of living organisms in terms of their dynamic, complex networks of metabolic, genetic, and epigenetic processes and signaling pathways. Alternative but closely related approaches focus on the interdependance of constraints, where constraints can be either molecular, such as enzymes, or macroscopic, such as the geometry of a bone or of the vascular system.
It has also been argued that the evolution of order in living systems and certain physical systems obeys a common fundamental principle termed the Darwinian dynamic. The Darwinian dynamic was formulated by first considering how macroscopic order is generated in a simple non-biological system far from thermodynamic equilibrium, and then extending consideration to short, replicating RNA molecules. The underlying order-generating process was concluded to be basically similar for both types of systems.
Another systemic definition called the operator theory proposes that "life is a general term for the presence of the typical closures found in organisms; the typical closures are a membrane and an autocatalytic set in the cell" and that an organism is any system with an organisation that complies with an operator type that is at least as complex as the cell. Life can also be modeled as a network of inferior negative feedbacks of regulatory mechanisms subordinated to a superior positive feedback formed by the potential of expansion and reproduction.
Some of the earliest theories of life were materialist, holding that all that exists is matter, and that life is merely a complex form or arrangement of matter. Empedocles (430 BC) argued that everything in the universe is made up of a combination of four eternal "elements" or "roots of all": earth, water, air, and fire. All change is explained by the arrangement and rearrangement of these four elements. The various forms of life are caused by an appropriate mixture of elements.
Democritus (460 BC) thought that the essential characteristic of life is having a soul (psyche). Like other ancient writers, he was attempting to explain what makes something a living thing. His explanation was that fiery atoms make a soul in exactly the same way atoms and void account for any other thing. He elaborates on fire because of the apparent connection between life and heat, and because fire moves.
Plato's world of eternal and unchanging Forms, imperfectly represented in matter by a divine Artisan, contrasts sharply with the various mechanistic Weltanschauungen, of which atomism was, by the fourth century at least, the most prominent ... This debate persisted throughout the ancient world. Atomistic mechanism got a shot in the arm from Epicurus ... while the Stoics adopted a divine teleology ... The choice seems simple: either show how a structured, regular world could arise out of undirected processes, or inject intelligence into the system.
— R.J. Hankinson, Cause and Explanation in Ancient Greek Thought
The mechanistic materialism that originated in ancient Greece was revived and revised by the French philosopher René Descartes, who held that animals and humans were assemblages of parts that together functioned as a machine. In the 19th century, the advances in cell theory in biological science encouraged this view. The evolutionary theory of Charles Darwin (1859) is a mechanistic explanation for the origin of species by means of natural selection.
Hylomorphism is a theory first expressed by the Greek philosopher Aristotle (322 BC). The application of hylomorphism to biology was important to Aristotle, and biology is extensively covered in his extant writings. In this view, everything in the material universe has both matter and form, and the form of a living thing is its soul (Greek psyche, Latin anima). There are three kinds of souls: the vegetative soul of plants, which causes them to grow and decay and nourish themselves, but does not cause motion and sensation; the animal soul, which causes animals to move and feel; and the rational soul, which is the source of consciousness and reasoning, which (Aristotle believed) is found only in man. Each higher soul has all of the attributes of the lower ones. Aristotle believed that while matter can exist without form, form cannot exist without matter, and that therefore the soul cannot exist without the body.
This account is consistent with teleological explanations of life, which account for phenomena in terms of purpose or goal-directedness. Thus, the whiteness of the polar bear's coat is explained by its purpose of camouflage. The direction of causality (from the future to the past) is in contradiction with the scientific evidence for natural selection, which explains the consequence in terms of a prior cause. Biological features are explained not by looking at future optimal results, but by looking at the past evolutionary history of a species, which led to the natural selection of the features in question.
Spontaneous generation was the belief that living organisms can form without descent from similar organisms. Typically, the idea was that certain forms such as fleas could arise from inanimate matter such as dust or the supposed seasonal generation of mice and insects from mud or garbage.
The theory of spontaneous generation was proposed by Aristotle,who compiled and expanded the work of prior natural philosophers and the various ancient explanations of the appearance of organisms; it held sway for two millennia. It was decisively dispelled by the experiments of Louis Pasteur in 1859, who expanded upon the investigations of predecessors such as Francesco Redi.Disproof of the traditional ideas of spontaneous generation is no longer controversial among biologists.
Vitalism is the belief that the life-principle is non-material. This originated with Georg Ernst Stahl (17th century), and remained popular until the middle of the 19th century. It appealed to philosophers such as Henri Bergson,Friedrich Nietzsche, and Wilhelm Dilthey, anatomists like Xavier Bichat, and chemists like Justus von Liebig. Vitalism included the idea that there was a fundamental difference between organic and inorganic material, and the belief that organic material can only be derived from living things. This was disproved in 1828, when Friedrich Wöhler prepared urea from inorganic materials. This Wöhler synthesis is considered the starting point of modern organic chemistry. It is of historical significance because for the first time an organic compound was produced in inorganic reactions.
During the 1850s, Hermann von Helmholtz, anticipated by Julius Robert von Mayer, demonstrated that no energy is lost in muscle movement, suggesting that there were no "vital forces" necessary to move a muscle. These results led to the abandonment of scientific interest in vitalistic theories, although the belief lingered on in pseudoscientific theories such as homeopathy, which interprets diseases and sickness as caused by disturbances in a hypothetical vital force or life force.
The age of the Earth is about 4.54 billion years. Evidence suggests that life on Earth has existed for at least 3.5 billion years, with the oldest physical traces of life dating back 3.7 billion years; however, some theories, such as the Late Heavy Bombardment theory, suggest that life on Earth may have started even earlier, as early as 4.1–4.4 billion years ago, and the chemistry leading to life may have begun shortly after the Big Bang,13.8 billion years ago, during an epoch when the universe was only 10–17 million years old.
More than 99% of all species of life forms, amounting to over five billion species, that ever lived on Earth are estimated to be extinct.
Although the number of Earth's catalogued species of lifeforms is between 1.2 million and 2 million, the total number of species in the planet is uncertain. Estimates range from 8 million to 100 million, with a more narrow range between 10 and 14 million, but it may be as high as 1 trillion (with only one-thousandth of one percent of the species described) according to studies realized in May 2016. The total number of related DNAbase pairs on Earth is estimated at 5.0 x 1037 and weighs 50 billion tonnes. In comparison, the total mass of the biosphere has been estimated to be as much as 4 TtC (trillion tons of carbon). In July 2016, scientists reported identifying a set of 355 genes from the Last Universal Common Ancestor (LUCA) of all organisms living on Earth.
All known life forms share fundamental molecular mechanisms, reflecting their common descent; based on these observations, hypotheses on the origin of life attempt to find a mechanism explaining the formation of a universal common ancestor, from simple organic molecules via pre-cellular life to protocells and metabolism. Models have been divided into "genes-first" and "metabolism-first" categories, but a recent trend is the emergence of hybrid models that combine both categories.
There is no current scientific consensus as to how life originated. However, most accepted scientific models build on the Miller–Urey experiment and the work of Sidney Fox, which show that conditions on the primitive Earth favored chemical reactions that synthesize amino acids and other organic compounds from inorganic precursors, and phospholipids spontaneously form lipid bilayers, the basic structure of a cell membrane.
Living organisms synthesize proteins, which are polymers of amino acids using instructions encoded by deoxyribonucleic acid (DNA). Protein synthesis entails intermediary ribonucleic acid (RNA) polymers. One possibility for how life began is that genes originated first, followed by proteins; the alternative being that proteins came first and then genes.
However, because genes and proteins are both required to produce the other, the problem of considering which came first is like that of the chicken or the egg. Most scientists have adopted the hypothesis that because of this, it is unlikely that genes and proteins arose independently.
Therefore, a possibility, first suggested by Francis Crick, is that the first life was based on RNA, which has the DNA-like properties of information storage and the catalytic properties of some proteins. This is called the RNA world hypothesis, and it is supported by the observation that many of the most critical components of cells (those that evolve the slowest) are composed mostly or entirely of RNA. Also, many critical cofactors (ATP,Acetyl-CoA,NADH, etc.) are either nucleotides or substances clearly related to them. The catalytic properties of RNA had not yet been demonstrated when the hypothesis was first proposed, but they were confirmed by Thomas Cech in 1986.
One issue with the RNA world hypothesis is that synthesis of RNA from simple inorganic precursors is more difficult than for other organic molecules. One reason for this is that RNA precursors are very stable and react with each other very slowly under ambient conditions, and it has also been proposed that living organisms consisted of other molecules before RNA. However, the successful synthesis of certain RNA molecules under the conditions that existed prior to life on Earth has been achieved by adding alternative precursors in a specified order with the precursor phosphate present throughout the reaction. This study makes the RNA world hypothesis more plausible.
In 2009, experiments demonstrated Darwinian evolution of a two-component system of RNA enzymes (ribozymes)in vitro. The work was performed in the laboratory of Gerald Joyce, who stated "This is the first example, outside of biology, of evolutionary adaptation in a molecular genetic system."
Prebiotic compounds may have originated extraterrestrially. NASA findings in 2011, based on studies with meteorites found on Earth, suggest DNA and RNA components (adenine,guanine and related organic molecules) may be formed in outer space.
The diversity of life on Earth is a result of the dynamic interplay between genetic opportunity, metabolic capability, environmental challenges, and symbiosis. For most of its existence, Earth's habitable environment has been dominated by microorganisms and subjected to their metabolism and evolution. As a consequence of these microbial activities, the physical-chemical environment on Earth has been changing on a geologic time scale, thereby affecting the path of evolution of subsequent life. For example, the release of molecular oxygen by cyanobacteria as a by-product of photosynthesis induced global changes in the Earth's environment. Because oxygen was toxic to most life on Earth at the time, this posed novel evolutionary challenges, and ultimately resulted in the formation of Earth's major animal and plant species. This interplay between organisms and their environment is an inherent feature of living systems.
The biosphere is the global sum of all ecosystems. It can also be termed as the zone of life on Earth, a closed system (apart from solar and cosmic radiation and heat from the interior of the Earth), and largely self-regulating. By the most general biophysiological definition, the biosphere is the global ecological system integrating all living beings and their relationships, including their interaction with the elements of the lithosphere,geosphere,hydrosphere, and atmosphere.
Life forms live in every part of the Earth's biosphere, including soil,hot springs,inside rocks at least 19 km (12 mi) deep underground, the deepest parts of the ocean, and at least 64 km (40 mi) high in the atmosphere. Under certain test conditions, life forms have been observed to thrive in the near-weightlessness of space and to survive in the vacuum of outer space. Life forms appear to thrive in the Mariana Trench, the deepest spot in the Earth's oceans. Other researchers reported related studies that life forms thrive inside rocks up to 580 m (1,900 ft; 0.36 mi) below the sea floor under 2,590 m (8,500 ft; 1.61 mi) of ocean off the coast of the northwestern United States, as well as 2,400 m (7,900 ft; 1.5 mi) beneath the seabed off Japan. In August 2014, scientists confirmed the existence of life forms living 800 m (2,600 ft; 0.50 mi) below the ice of Antarctica. According to one researcher, "You can find microbes everywhere—they're extremely adaptable to conditions, and survive wherever they are."
In a general sense, biospheres are any closed, self-regulating systems containing ecosystems. This includes artificial biospheres such as Biosphere 2 and BIOS-3, and potentially ones on other planets or moons.
The inert components of an ecosystem are the physical and chemical factors necessary for life—energy (sunlight or chemical energy), water, heat, atmosphere,gravity,nutrients, and ultravioletsolar radiation protection. In most ecosystems, the conditions vary during the day and from one season to the next. To live in most ecosystems, then, organisms must be able to survive a range of conditions, called the "range of tolerance." Outside that are the "zones of physiological stress," where the survival and reproduction are possible but not optimal. Beyond these zones are the "zones of intolerance," where survival and reproduction of that organism is unlikely or impossible. Organisms that have a wide range of tolerance are more widely distributed than organisms with a narrow range of tolerance.
To survive, selected microorganisms can assume forms that enable them to withstand freezing,complete desiccation,starvation, high levels of radiation exposure, and other physical or chemical challenges. These microorganisms may survive exposure to such conditions for weeks, months, years, or even centuries. Extremophiles are microbial life forms that thrive outside the ranges where life is commonly found. They excel at exploiting uncommon sources of energy. While all organisms are composed of nearly identical molecules, evolution has enabled such microbes to cope with this wide range of physical and chemical conditions. Characterization of the structure and metabolic diversity of microbial communities in such extreme environments is ongoing.
Microbial life forms thrive even in the Mariana Trench, the deepest spot in the Earth's oceans. Microbes also thrive inside rocks up to 1,900 feet (580 m) below the sea floor under 8,500 feet (2,600 m) of ocean.
Investigation of the tenacity and versatility of life on Earth, as well as an understanding of the molecular systems that some organisms utilize to survive such extremes, is important for the search for life beyond Earth. For example, lichen could survive for a month in a simulated Martian environment.
All life forms require certain core chemical elements needed for biochemical functioning. These include carbon,hydrogen,nitrogen,oxygen,phosphorus, and sulfur—the elemental macronutrients for all organisms — often represented by the acronym CHNOPS. Together these make up nucleic acids, proteins and lipids, the bulk of living matter. Five of these six elements comprise the chemical components of DNA, the exception being sulfur. The latter is a component of the amino acids cysteine and methionine. The most biologically abundant of these elements is carbon, which has the desirable attribute of forming multiple, stable covalent bonds. This allows carbon-based (organic) molecules to form an immense variety of chemical arrangements. Alternative hypothetical types of biochemistry have been proposed that eliminate one or more of these elements, swap out an element for one not on the list, or change required chiralities or other chemical properties.
DNA stores biological information. The DNA backbone is resistant to cleavage, and both strands of the double-stranded structure store the same biological information. Biological information is replicated as the two strands are separated. A significant portion of DNA (more than 98% for humans) is non-coding, meaning that these sections do not serve as patterns for protein sequences.
The two strands of DNA run in opposite directions to each other and are therefore anti-parallel. Attached to each sugar is one of four types of nucleobases (informally, bases). It is the sequence of these four nucleobases along the backbone that encodes biological information. Under the genetic code,RNA strands are translated to specify the sequence of amino acids within proteins. These RNA strands are initially created using DNA strands as a template in a process called transcription.
Within cells, DNA is organized into long structures called chromosomes. During cell division these chromosomes are duplicated in the process of DNA replication, providing each cell its own complete set of chromosomes. Eukaryotic organisms (animals, plants, fungi, and protists) store most of their DNA inside the cell nucleus and some of their DNA in organelles, such as mitochondria or chloroplasts. In contrast, prokaryotes (bacteria and archaea) store their DNA only in the cytoplasm. Within the chromosomes, chromatin proteins such as histones compact and organize DNA. These compact structures guide the interactions between DNA and other proteins, helping control which parts of the DNA are transcribed.
The first known attempt to classify organisms was conducted by the Greek philosopher Aristotle (384–322 BC), who classified all living organisms known at that time as either a plant or an animal, based mainly on their ability to move. He also distinguished animals with blood from animals without blood (or at least without red blood), which can be compared with the concepts of vertebrates and invertebrates respectively, and divided the blooded animals into five groups: viviparous quadrupeds (mammals), oviparous quadrupeds (reptiles and amphibians), birds, fishes and whales. The bloodless animals were also divided into five groups: cephalopods,crustaceans, insects (which included the spiders, scorpions, and centipedes, in addition to what we define as insects today), shelled animals (such as most molluscs and echinoderms), and "zoophytes" (animals that resemble plants). Though Aristotle's work in zoology was not without errors, it was the grandest biological synthesis of the time and remained the ultimate authority for many centuries after his death.
The exploration of the Americas revealed large numbers of new plants and animals that needed descriptions and classification. In the latter part of the 16th century and the beginning of the 17th, careful study of animals commenced and was gradually extended until it formed a sufficient body of knowledge to serve as an anatomical basis for classification.
In the late 1740s, Carl Linnaeus introduced his system of binomial nomenclature for the classification of species. Linnaeus attempted to improve the composition and reduce the length of the previously used many-worded names by abolishing unnecessary rhetoric, introducing new descriptive terms and precisely defining their meaning. The Linnaean classification has eight levels: domains, kingdoms, phyla, class, order, family, genus, and species.
The fungi were originally treated as plants. For a short period Linnaeus had classified them in the taxon Vermes in Animalia, but later placed them back in Plantae. Copeland classified the Fungi in his Protoctista, thus partially avoiding the problem but acknowledging their special status. The problem was eventually solved by Whittaker, when he gave them their own kingdom in his five-kingdom system.Evolutionary history shows that the fungi are more closely related to animals than to plants.
As new discoveries enabled detailed study of cells and microorganisms, new groups of life were revealed, and the fields of cell biology and microbiology were created. These new organisms were originally described separately in protozoa as animals and protophyta/thallophyta as plants, but were united by Haeckel in the kingdom Protista; later, the prokaryotes were split off in the kingdom Monera, which would eventually be divided into two separate groups, the Bacteria and the Archaea. This led to the six-kingdom system and eventually to the current three-domain system, which is based on evolutionary relationships. However, the classification of eukaryotes, especially of protists, is still controversial.
As microbiology, molecular biology and virology developed, non-cellular reproducing agents were discovered, such as viruses and viroids. Whether these are considered alive has been a matter of debate; viruses lack characteristics of life such as cell membranes, metabolism and the ability to grow or respond to their environments. Viruses can still be classed into "species" based on their biology and genetics, but many aspects of such a classification remain controversial.
In May 2016, scientists reported that 1 trillion species are estimated to be on Earth currently with only one-thousandth of one percent described.
The original Linnaean system has been modified over time as follows:
Cells are the basic unit of structure in every living thing, and all cells arise from pre-existing cells by division.Cell theory was formulated by Henri Dutrochet,Theodor Schwann,Rudolf Virchow and others during the early nineteenth century, and subsequently became widely accepted. The activity of an organism depends on the total activity of its cells, with energy flow occurring within and between them. Cells contain hereditary information that is carried forward as a genetic code during cell division.
There are two primary types of cells. Prokaryotes lack a nucleus and other membrane-bound organelles, although they have circular DNA and ribosomes. Bacteria and Archaea are two domains of prokaryotes. The other primary type of cells are the eukaryotes, which have distinct nuclei bound by a nuclear membrane and membrane-bound organelles, including mitochondria,chloroplasts,lysosomes, rough and smooth endoplasmic reticulum, and vacuoles. In addition, they possess organized chromosomes that store genetic material. All species of large complex organisms are eukaryotes, including animals, plants and fungi, though most species of eukaryote are protistmicroorganisms. The conventional model is that eukaryotes evolved from prokaryotes, with the main organelles of the eukaryotes forming through endosymbiosis between bacteria and the progenitor eukaryotic cell.
The molecular mechanisms of cell biology are based on proteins. Most of these are synthesized by the ribosomes through an enzyme-catalyzed process called protein biosynthesis. A sequence of amino acids is assembled and joined together based upon gene expression of the cell's nucleic acid. In eukaryotic cells, these proteins may then be transported and processed through the Golgi apparatus in preparation for dispatch to their destination.
Cells reproduce through a process of cell division in which the parent cell divides into two or more daughter cells. For prokaryotes, cell division occurs through a process of fission in which the DNA is replicated, then the two copies are attached to parts of the cell membrane. In eukaryotes, a more complex process of mitosis is followed. However, the end result is the same; the resulting cell copies are identical to each other and to the original cell (except for mutations), and both are capable of further division following an interphase period.
Multicellular organisms may have first evolved through the formation of colonies of identical cells. These cells can form group organisms through cell adhesion. The individual members of a colony are capable of surviving on their own, whereas the members of a true multi-cellular organism have developed specializations, making them dependent on the remainder of the organism for survival. Such organisms are formed clonally or from a single germ cell that is capable of forming the various specialized cells that form the adult organism. This specialization allows multicellular organisms to exploit resources more efficiently than single cells. In January 2016, scientists reported that, about 800 million years ago, a minor genetic change in a single molecule, called GK-PID, may have allowed organisms to go from a single cell organism to one of many cells.
Cells have evolved methods to perceive and respond to their microenvironment, thereby enhancing their adaptability. Cell signaling coordinates cellular activities, and hence governs the basic functions of multicellular organisms. Signaling between cells can occur through direct cell contact using juxtacrine signalling, or indirectly through the exchange of agents as in the endocrine system. In more complex organisms, coordination of activities can occur through a dedicated nervous system.
Though life is confirmed only on Earth, many think that extraterrestrial life is not only plausible, but probable or inevitable. Other planets and moons in the Solar System and other planetary systems are being examined for evidence of having once supported simple life, and projects such as SETI are trying to detect radio transmissions from possible alien civilizations. Other locations within the Solar System that may host microbial life include the subsurface of Mars, the upper atmosphere of Venus, and subsurface oceans on some of the moons of the giant planets. Beyond the Solar System, the region around another main-sequence star that could support Earth-like life on an Earth-like planet is known as the habitable zone. The inner and outer radii of this zone vary with the luminosity of the star, as does the time interval during which the zone survives. Stars more massive than the Sun have a larger habitable zone, but remain on the Sun-like "main sequence" of stellar evolution for a shorter time interval. Small red dwarfs have the opposite problem, with a smaller habitable zone that is subject to higher levels of magnetic activity and the effects of tidal locking from close orbits. Hence, stars in the intermediate mass range such as the Sun may have a greater likelihood for Earth-like life to develop. The location of the star within a galaxy may also affect the likelihood of life forming. Stars in regions with a greater abundance of heavier elements that can form planets, in combination with a low rate of potentially habitat-damaging supernova events, are predicted to have a higher probability of hosting planets with complex life. The variables of the Drake equation are used to discuss the conditions in planetary systems where civilization is most likely to exist. Use of the equation to predict the amount of extraterrestrial life, however, is difficult; because many of the variables are unknown, the equation functions as more of a mirror to what its user already thinks. As a result, the number of civilizations in the galaxy can be estimated as low as 9.1 x 10−13, suggesting a minimum value of 1, or as high as 15.6 million (0.156 x 109); for the calculations, see Drake equation.
Artificial life is the simulation of any aspect of life, as through computers, robotics, or biochemistry. The study of artificial life imitates traditional biology by recreating some aspects of biological phenomena. Scientists study the logic of living systems by creating artificial environments—seeking to understand the complex information processing that defines such systems. While life is, by definition, alive, artificial life is generally referred to as data confined to a digital environment and existence.
Synthetic biology is a new area of biotechnology that combines science and biological engineering. The common goal is the design and construction of new biological functions and systems not found in nature. Synthetic biology includes the broad redefinition and expansion of biotechnology, with the ultimate goals of being able to design and build engineered biological systems that process information, manipulate chemicals, fabricate materials and structures, produce energy, provide food, and maintain and enhance human health and the environment.
One of the challenges in defining death is in distinguishing it from life. Death would seem to refer to either the moment life ends, or when the state that follows life begins.However, determining when death has occurred is difficult, as cessation of life functions is often not simultaneous across organ systems. Such determination therefore requires drawing conceptual lines between life and death. This is problematic, however, because there is little consensus over how to define life. The nature of death has for millennia been a central concern of the world's religious traditions and of philosophical inquiry. Many religions maintain faith in either a kind of afterlife or reincarnation for the soul, or resurrection of the body at a later date.
Extinction is the process by which a group of taxa or species dies out, reducing biodiversity. The moment of extinction is generally considered the death of the last individual of that species. Because a species' potential range may be very large, determining this moment is difficult, and is usually done retrospectively after a period of apparent absence. Species become extinct when they are no longer able to survive in changing habitat or against superior competition. In Earth's history, over 99% of all the species that have ever lived are extinct; however, mass extinctions may have accelerated evolution by providing opportunities for new groups of organisms to diversify.
Fossils are the preserved remains or traces of animals, plants, and other organisms from the remote past. The totality of fossils, both discovered and undiscovered, and their placement in fossil-containing rock formations and sedimentary layers (strata) is known as the fossil record. A preserved specimen is called a fossil if it is older than the arbitrary date of 10,000 years ago. Hence, fossils range in age from the youngest at the start of the Holocene Epoch to the oldest from the Archaean Eon, up to 3.4 billion years old.